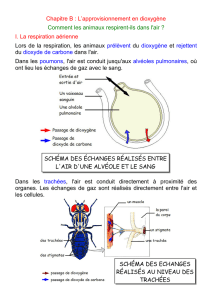
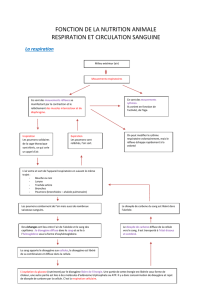
La respiration chez les animaux

perso.ens-lyon.org/romain.berardozzi
La respiration chez les animaux
Introduction et problématique
I. La respiration: un échange de gaz par diffusion
II. Contraintes du milieu aérien et du milieu aquatique
III. Les appareils respiratoires en milieu aquatique
IV. Les appareils respiratoires en milieu aérien
V. Modulation du fonctionnement des appareils respiratoires
VI. Origine et évolution des appareils respiratoires
Conclusion / Bilan
Introduction
Les animaux (métazoaires) sont des organismes pluricellulaires eucaryotes pouvant être
considérés comme des machines thermodynamiques ouvertes sur leur environnement avec
lequel ils échangent de la matière et de l'énergie. Afin de répondre à leurs besoins
énergétiques (s'élevant à 2800 kcal par jour pour un Homme male adulte de 70 kg exerçant
une activité physique modérée), ils puisent dans leur environnement les molécules
organiques qu'ils oxydent pour aboutir à la formation de molécules d'ATP (environ 65 kg par
jour pour satisfaire aux besoins énergétiques), énergie directement utilisable par la cellule
pour son fonctionnement.
En présence de dioxygène (aérobiose), les cellules oxydent totalement les
molécules organiques par respiration cellulaire (glycolyse cytoplasmique + cycle de
Krebs et chaine respiratoire mitochondriale) ce qui permet la formation de 36 ATP (2
par glycolyse et 34 par la chaine respiratoire + cycle de Krebs). Pour le glucose, la
réaction est la suivante:
C6H12O6+6 O2+36 ATP +36 Pi → 6 CO2+6 H2O+ 36 ATP
En l'absence de dioxygène (anaérobiose), les cellules oxydent partiellement les
molécules organiques par fermentation (glycolyse et fermentation lactique
cytoplasmiques) ce qui permet la formation de 2 ATP seulement.

perso.ens-lyon.org/romain.berardozzi
En conséquence, l'oxydation des molécules organiques par respiration est 19 fois plus
efficace que l'oxydation par fermentation en terme de production d'ATP par mole de
glucose oxydé. Aussi, l'approvisionnement en dioxygène de chaque cellule de l'organisme
pluricellulaire d'un animal autorise de grandes économies et permet de couvrir des besoins
énergétiques plus élevés.
D'autre part, la respiration cellulaire conduit à l'accumulation de déchets métaboliques dont
le dioxyde de carbone. La réaction du dioxyde de carbone avec l'eau conduit à l'acidification
des cellules selon la réaction suivante: CO2+H2O=HCO3-+H+. L'acidification peut entrainer
une altération de l'activité enzymatique due à la modification des structures
tridimensionnelles. Une mauvaise évacuation des déchets métaboliques entraine en
conséquent des effets délétères pour la cellule et l'animal.
Comment les organismes prélèvent-ils le dioxygène nécessaire à la couverture de
leur besoins énergétiques? Par quelles modalités? Au niveau de quelles
structures?
Comment le dioxygène est-il véhiculé jusque chaque cellule? Comment les
déchets sont-ils évacués?
Les animaux vivent dans des milieux aux caractéristiques différentes: eau ou air. Au cours
de leur développement embryonnaire et post-embryonnaire, les animaux peuvent évoluer
dans des milieux différents. De plus, au cours de leur vie, les animaux peuvent être soumis à
des variations temporaires des conditions physico-chimiques du milieu.
En quoi ces structures sont-elles adaptées au milieu de vie de l'animal?
Comment ces structures sont en mesure de s'adapter temporairement aux
contraintes du milieu de vie et aux contraintes métaboliques internes (exercice
physique et demande en ATP notamment)?
D'où viennent ces structures et comment ont-elles évoluées au cours des temps
géologiques?

perso.ens-lyon.org/romain.berardozzi
La respiration: un échange de gaz par diffusion
entre milieu de vie et milieu intérieur
La respiration cellulaire et métabolisme
Comme il a été souligné dans l'introduction, le rendement de la respiration est 19 fois
supérieur à celui de la fermentation en terme de production d'ATP. L'absence
d'approvisionnement des cellules en dioxygène (ou un approvisionnement trop faible) peut
avoir des effets graves pour l'animal. Chez l'Homme, l'anoxie des cellules du cerveau suite à
un accident vasculaire cérébral par exemple, entraine dans les premières secondes des
troubles du langage, de la vue, de la pensée dûs à l'arrêt du fonctionnement cellulaire. Si
l'anoxie se prolonge, les tissus commencent à subir des effets irréversibles liés à la mort des
cellules par nécrose. Plus de 5 minutes d'anoxie des cellules du cerveau conduit à la mort
cérébral. Chez l'Homme, le dioxygène est nécessaire à la survie.
D'où vient ce dioxygène nécessaire à la survie de l'organisme?
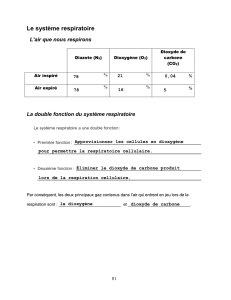
Les animaux échangent des gaz respiratoires avec leur milieu de vie
La composition de l'atmosphère révèle la présence de dioxygène (~21%) et de dioxyde de
carbone (~0.03%) entre autres. Les expériences historiques de Lavoisier ont permis de
mettre en évidence que les animaux prélèvent dans l'atmosphère du dioxygène (en
atmosphère confinée cela fini par éteindre les bougies par absence de comburant) et
relargue du dioxyde de carbone (en atmosphère confinée cela fini par troubler l'eau de
chaux).
« J’ai mis un moineau sous une cloche de verre remplie d’air : l’animal n’a paru nullement
affecté dans les premiers instants ; au bout d’un quart d’heure, il a commencé à s’agiter, sa
respiration est devenue précipitée et pénible. Cet air, qui avait été respiré par l’animal, était
devenu fort différent de l’air de l’atmosphère ; il précipitait l’eau de chaux ; il éteignait les
bougies » Antoine Lavoisier
Après cette expérience, Lavoisier émit 2 hypothèses quant au devenir des gaz respiratoires:
le dioxygène entre au niveau des poumons et y est transformé localement en dioxyde
de carbone: les poumons sont les organes de la combustion.
le dioxygène entre au niveau des poumons puis est distribué à l'ensemble de
l'organisme où la combustion à lieu afin de produire du dioxyde de carbone ensuite
évacué au niveau des poumons
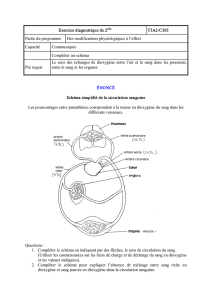
L'étude des sangs artériels et veineux au niveau des poumons permet de montrer que les
gaz respiratoires ne restent pas au niveau des poumons mais passent dans le milieu
intérieur via la circulation sanguine au niveau des capillaires pulmonaires.

perso.ens-lyon.org/romain.berardozzi
Sang artériel pulmonaire
Sang veineux artériel
Dioxyde de carbone
6.13 kPa
50 mmHg
560 mL/L
5.33 kPa
40 mmHg
520 mL/L
Dioxygène
5.33 kPa
40 mmHg
160 mL/L
13.33 kPa
100 mmHg
200 mL/L
L'expérience de Lavoisier peut être réalisée de manière plus moderne en mesurant les
concentrations en dioxygène et en dioxyde de carbone de l'air confiné sous cloche dans
lequel on place un criquet par exemple. L'air sous la cloche au cours du temps est déplété
en dioxygène et se concentre en dioxyde de carbone.
Au niveau de quelles structures et selon quelles modalités les gaz respiratoires
passent-ils du milieu de vie vers le milieu intérieur et inversement?
Le transport des gaz respiratoires au niveau de l'interface milieu de vie / milieu
intérieur se fait par diffusion et est régi par la loi de Fick
La diffusion est un phénomène physique de transport irréversible et sans coût énergétique
des molécules de proche en proche des compartiments les plus concentrés vers les
compartiments les moins concentrés. Le flux de diffusion (noté J) exprimé en mol.s-1 ou g.s-
1 est donné par la loi de Fick.
J=Kgaz . S/e .ΔPgaz
avec Kgaz le coefficient de diffusion du gaz (dioxygène ou dioxyde de carbone) exprimée en m2.s-1, S
la surface de l'interface, e l'épaisseur de l'interface et ΔPgaz la différence de pression partielle en gaz
de part et d'autre de l'interface.
Ce phénomène physique de transport présente l'avantage de ne pas avoir de coût
énergétique pour la cellule. En revanche, le transport se faisant de proche en proche limite la
distance que peuvent parcourir les gaz.
L'échange de gaz par diffusion est autorisé par la perméabilité des membranes
biologiques aux petites molécules gazeuses (dioxygène et dioxyde de carbone notamment).

perso.ens-lyon.org/romain.berardozzi
La loi de Harvey illustre ce propos en indiquant qu'en milieu aquatique, toutes les
cellules d'un organisme sphérique peuvent être approvisionnées en dioxygène si et
seulement si son diamètre ne dépasse pas 2 mm.
Or, en milieu aquatique, de nombreux animaux ont une taille qui dépasse largement
ces 2 mm de diamètre. Aussi, afin de contourner les limites imposées par la courte
distance de diffusion des gaz, certains organismes voient la forme de leur corps
optimiser le rapport surface sur volume (S/V) afin que toutes les cellules du corps
(même les plus distantes de l'interface milieu de vie / milieu intérieur) soient séparées
d'une distance inférieure à la distance maximale sur laquelle la diffusion est possible.
Les formes permettant d'avoir un rapport S/V le plus grand (donc le plus en faveur
d'une diminution de la distance interface / cellules profondes) sont les formes
allongées (vers) et plates (plathelminthes, cténophore par exemple). Au contraire, les
formes sphériques ou cubiques ont un rapport S/V défavorable pour
l'approvisionnement par diffusion seule. Dans ce cas-là, le transport des gaz
respiratoires se fait par un système de transport de matière par convection: c'est la
circulation des fluides intérieurs (sang ou hémolymphe).
Sur le schéma suivant, les cellules pour lesquelles l'approvisionnement en gaz n'est
pas possible par diffusion sont représentées en blanc. On remarque que pour les
formes plates et allongées, ces cellules sont très peu abondantes et la diffusion
permet d'approvisionner quasiment toutes les cellules de l'organisme.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
1
/
24
100%