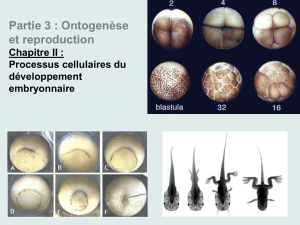
Aucun titre de diapositive - bcpst-svt-parc

Chapitre I :
Mise en place du plan d ’organisation de la Grenouille :
de la cellule-œuf à l ’organisme adulte

La polarité de la cellule-œuf héritée de l ’ovocyte II
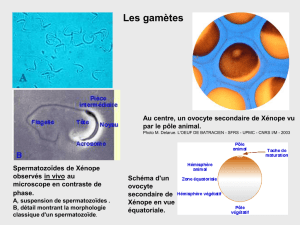
Au centre, un ovocyte II de
Xénope vu par le pôle animal.
Photo M. Delarue. L'OEUF DE BATRACIEN - SFRS -
UPMC - CNRS I/M - 2003
Document 1. L’œuf de Grenouille en
vue latérale gauche.
[PATTIER J.-Y., “ Croissance et développement des animaux ”, Ellipses
Ed., 1991].
Ovocyte II de Xénope en vue équatoriale.

Après avoir traversé les
gangues entourant l'ovocyte,
le spermatozoïde perce la
« membrane » vitelline puis
vient au contact de la
membrane plasmique (A à D).
La fusion du spermatozoide
et de l'ovocyte entraîne une
série de réactions
cytoplasmiques et
membranaires dont la plus
visible est le soulèvement de
la « membrane » vitelline en
une « membrane » de
fécondation (D et E).
L'espace contenu entre la
membrane plasmique et la
membrane de fécondation
porte le nom d'espace
périvitellin (E).
Les enveloppes protectrices
de l’ovocyte II puis de l’œuf

Répartition des ARN
(coloration par le bleu de toluidine)
Les ARN sont bien plus nombreux
dans l'hémisphère animal. La majeure
partie des ARN cytoplasmiques étant
ribosomiques, la coloration reflète le
gradient ribosomique.
Coupe méridienne d'ovocyte II
de pleurodèle
On observe une nette différence
de charge en vitellus entre le pôle
animal et le pôle végétatif.
L’ovocyte secondaire est polarisé

L’ovogenèse chez la Grenouille
Croissance et
accumulation de réserves
(3e année d’ovogenèse)
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
1
/
86
100%