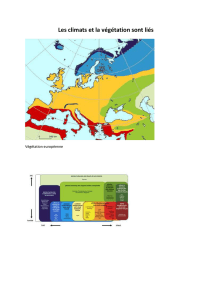
Ecologie et dynamique de la végétation de l`Aurès

REPUBLIQUE ALGERIENNE DEMOCRATIQUE ET POPULAIRE
MINISTERE DE L'ENSEIGNEMENT SUPERIEUR ET DE LA
RECHERCHE SCIENTIFIQUE
Université Mohamed Khider – Biskra -
Faculté des sciences exactes et des sciences de la vie
Département d'agronomie
Présentée par
Mr. Yassine BEGHAMI
Pour l’obtention du diplôme de doctorat en sciences agronomiques
Thème
Ecologie et dynamique de la
végétation de l’A urès :
analyse spatio-temporelle et étude de
la flore forestière et montagnarde
Soutenue publiqueme nt le : 8 janvier 2013
Devant le jury composé de :
Président : Mr. Mohamed Belhamra : Pr. Université de Biskra -
Directeur de thèse : Mr. Mahdi Kalla : Pr. Université de Batna -
Codirecteur : Mr. Thinon Michel : chargé de recherche, CNR S Marseille, France
Examinateur : Mr. Rabeh Bensaid : Pr. Université de Skikda –
Examinateur : Mr. Djamel Allatou : Pr. Université de Constantine –
Année universitaire 2011 -2012
Thèse

Dédicace
Je dédie ce précieux travail aux personnes qui me sont les plus chères au
monde.
A la mémoire de mes grands parents Amor et M’louka que Dieu Tout
Puissant les accueille dans son vaste paradis .
A mes chers parents Messaoud et Zohra pour tous leur s sacrifices et leurs
dévouement à mon égard .
A ma femme Akila qui s’est sacrifiée à mes étude s depuis mon magistère et
mon doctorat. La femme qui a su me soutenir moralement et
matériellement durant les pires moments de mes aventures sur le terrain, le
traitement de mes échantillons et la rédaction de ce manuscrit.
A mes chers gosses Massil, Mazigh et Maria, à qui j’ai probablement
manqué un peu de tendresse et d’affection. Ils n’ont jamais osé de tâter le
désordre de ma chambre en particulier les feuilles et les plantes de mon
herbier éparpillées par tout .
A mes frères et sœurs : Mohamed, Jamel, Zineb, Rachid, Haba, Akila et
Ammar
A mes amis Mehdi et Gaga
A tous les amoureux d’une nature propre et saine
Je dédie cette modeste recherche
Yassine (Aurès)

Remerciements
Au terme de ce travail, il m’est particulièrement agréable de remercier toutes
personnes ayant contribué de prés ou de loin à la réalisation de cette modeste
recherche.
Tous mes remerciements vont d’abords à Monsieur le Professeur mahdi Ka lla,
directeur du laboratoire LRNAT de l’université de Batna, pour avoir accepté de
diriger ce travail, qu’il trouve ici, l’expression de ma profonde et sincère
reconnaissance, pour tous ses efforts fourni à mon égard le long de ce travail , son
savoir, ses critiques constructives, et sa confiance étaient pour moi d’un grand
secours.
Aucun remerciement ne saurait être à la hauteur de toute l’aide et de tout le soutien
que Michel Thinon , directeur de recherche au CNRS, m’a apportés en co -dirigeant
cette thèse. Il m’a accueilli dans son laboratoire de pédoanthracologie à l’IMEP,
Université Paul Cézanne Marseille III. Grâce à son créateur (Michel Thinon), j’ai
appris une nouvelle technique d’étude de la dynamique de la végétation. Mes séjours
au laboratoire de p édoanthracologie m’ont permis de découvrir une nouvelle approche
de la dynamique des écosystèmes Méditerranéens en visitant particulièrement la forêt
de Sainte baume. Je lui en serai toujours reconnaissante.
Ma très vive gratitude va aussi à monsieur Belh amra Mohamed professeur à
l’université Mohamed Kheider de Biskra de m’avoir fait l’honneur d’accepter de
présider le jury de ma soutenance
Mes sincères remerciements vont à Mr le Professeur Djamel Allatou Professeur à la
Faculté des sciences de la nature et de la vie, université Mentouri Constantine de
m’avoir fait l’honneur d’examiner mon travail. Qu’il trouve ici tous les respects les
plus sincères d’élève à son professeur.
Il m’est agréable aussi d’adresser un grand merci pour notre cher professeur Rabeh
Bensaid de l’université de Skikda qui a bien voulu prendre part dans le jury et
examiner mon travail. Il trouve ici mes sincères remerciements.

Monsieur Errol Véla , cher ami, et maître de conférence à l’université de Montpellier
qui, grâce à son aide et son soutien, ce travail aurait pu voir le jour. Malgré, ces
nombreuses tâche et fonctions, il n’a pas ménagé ses efforts pour m’encourager, et
m’aider. En tant que spécialiste de la flore de l’Afrique du nord, il a manifesté un
intérêt particulier pour ce travail. Qu’il trouve ici l’expression de ma vive gratitude
et reconnaissance.
La rencontre avec le professeur Gérard de Bélair a été sans doute l’un des
déterminants essentiels pour mon activité de recherche suivante. Je tiens à le
remercier du fond du cœur pour la confiance qu’il m’a accordé, l’intérêt qu’il a porté
à mon égard ainsi qu’à mon travail et surtout son aide précieuse de laquelle j’ai
bénéficié pendant les moments difficiles. Qu’il trouve ici ma sincère gratitude
De nombreuses personne s ont régulièrement contribué à la réalisation de cette
recherche, je dois citer ici particulièrement les services forestiers des conservations de
Btana et Khenchela. Mes cordiaux remerciement s vont aux messieurs : Briki
Othman, Bouhrik, Boumaaraf, Bouzgha ya A, Zekak S, Hammedi, Aksal R, Laassis
et Kalil N, Abida Omar, Minasri A, Iouadj A avec lesquels j’ai passé de bons
moments en pleine nature et pour le ur soutien logistique et assistance qu’ils ont
assurés.
De nombreux collègues et amis qui ont voulu s à chaque fois réenchanter la science
pendant cette thèse. Parmi lesquels, citons Abdessalem S, Benmessaoud H , Djerah A.
et Aboubou Abbes Qu’ils trouvent ici mes vives gratitudes
Mes remerciements von t également à mes étudiants, que j’ai encadré pour soute nir
leur ingéniorats. Leurs présences sur le terrain et leurs accompagnements étaient ,
pour moi, d’un grand secours en stimulant mon engouement à découvrir les coins les
plus isolés et reculés des Aurès. Je cite ici Guermat R, Deouak H et Zaatour S.

Sommaire
Page
Introduction générale
01
Partie I : Présentation de la zone d’étude
Chapitre 1 : Cadre physique
I. 1.
Localisation géographique
05
I. 2.
Situation administrative
05
I. 3.
Géographie et limites de l'Aurès
07
I. 4.
Hydrographie
09
I. 5.
Les grands traits de la paléogéographie de l’Aurès
10
I. 6.
Sol
10
I. 7.
Géologie
12
Chapitre 2 : Cadre climatique et bioclimatique
I. 2.
Conditions climatiques et bioclimatiques
15
Introduction
15
I. 2. 1
Matériel et méthodes
16
I. 2. 2.
Paramètres climatiques
16
I. 2. 2. 1.
Les précipitations
16
I. 2. 2. 2.
Températures
24
I. 2. 2. 3.
Les gradients
27
Conclusion
42
Partie II : analyse écologique et biogéographique
Chapitre 1 : Aspects méthodologiques généraux
II. 1.
Aspects méthodologiques généraux
43
Introduction
43
II. 1. 1.
Choix de la zone d’étude
43
II. 1. 2.
Stratification de l’espace
43
II. 1. 3.
Organisation du plan d’échantillonnage
44
II. 1. 4.
Réalisation du relevé et collecte des données
47
Chapitre 2 : Intérêt de la diversité floristique
II. 2.
Intérêt de la diversité floristique de la cédraie des Ouled Yagoub
49
Introduction
49
II. 2. 1.
Site d’étude et méthodologie
50
II. 2. 2.
Echantillonnage
52
II. 2. 3.
Résultats et discussions
52
Conclusion
61
Chapitre 3 : Les formations arborées pré-steppiques d’altitude
II. 3.
Etude des formations arborées pré-steppiques d’altitude
63
Introduction
63
II. 3. 1.
Site et Méthodes
63
II. 3. 2.
Résultats et discussions
65
Conclusion
74
II. 4.
Chapitre 4 : La végétation du massif de Chélia
La végétation du massif du Chélia : Typologie et signification
écologique.
76
Introduction
76
II. 4. 1.
Matériel et Méthodes
76
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
231
232
233
234
235
236
237
238
239
240
241
242
243
244
245
246
247
248
249
250
251
252
253
254
255
256
257
258
259
260
261
262
263
264
265
266
267
268
269
270
271
272
273
274
275
276
277
278
279
280
281
282
283
284
285
286
287
288
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
231
232
233
234
235
236
237
238
239
240
241
242
243
244
245
246
247
248
249
250
251
252
253
254
255
256
257
258
259
260
261
262
263
264
265
266
267
268
269
270
271
272
273
274
275
276
277
278
279
280
281
282
283
284
285
286
287
288
1
/
288
100%