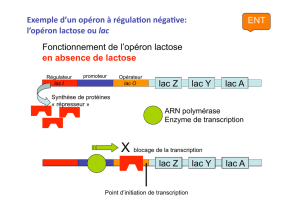
Opérateur lac

Cours
Module L5-BH-05
Régulation de l’expression des génomes
Mercredi de 13h30 à 15h30 & Jeudi de 8h00 à 10h00 (sem. 0 à 5/6)
Université d’Orléans – UFR Sciences & Centre de Biophysique Moléculaire UPR4301
S. Bourgerie
*1

PLAN des COURS n°1 & n°2
1- Introduction
1.1 pourquoi y-a-t-il régulation?
1.2 niveaux de régulation
1.3 quelques définitions
1.4 notion d’opéron
2- Régulation de la transcription
2.1 stratégies de contrôle de l’initiation de la transcription
2.2 contrôles positif-négatif
2.3 rappels sur l’initiation de la transcription chez les procaryotes
3- Opéron lactose
3.1 organisation générale
3.2 phénomène d’induction - inducteur
3.3 lacZ - lacY - lacA
3.4 mutants de l’opéron
3.5 régulations
3.5.1 négative (répresseur)
3.5.1.1 expérience Pajamo
3.5.1.2 rôle du répresseur
3.5.1.3 structure du répresseur
3.5.1.4 opérateur
3.5.2 positive : répression catabolique
3.5.2.1 diauxie
3.5.2.2 AMPc
3.5.2.3 scénarios
3.5.2.4 structure-fonction CRP
3.5.2.5 opérateurs
3.6 récapitulatif
*2

Introduction Pourquoi y-a-t-il régulation de l’expression des gènes ?
Pour répondre aux conditions changeantes
de l’environnement immédiat
Sept mécanismes susceptibles de modifier la concentration à l’équilibre d’une protéine ;
Sites possibles de régulation :
1- synthèse du transcrit primaire
2- maturation post-transcriptionnelle de l’ARNm
3- dégradation de l’ARNm
4- synthèse protéique
5- modifications post-traductionnelles
6- ciblage et transport des protéines
7- dégradation des protéines
*3

1
23
4
5
6
7
*4

Quelques définitions
Les séquences codant des produits agissant en trans
Les séquences agissant en cis
Un locus agit en cis sur un autre locus s’il est sur la même molécule d’ADN ;
L’opérateur est un élément d’activation en cis car il fonctionne uniquement
s’il est fixé sur le gène dont il contrôle l’expression.
Un locus agit en trans s’il peut contrôler un autre locus, même à partir d’une molécule
d’ADN différente.
Le gène du répresseur du lactose (lacI) agit en trans car il peut réguler l’expression
de l’opéron lactose même s’il est enlevé du génome d’E. coli et placé sur un plasmide.
5
élément régulateur en cis : site pour une protéine de liaison à l’ADN, situé le plus souvent,
en amont du gène régulé.
élément agissant en trans : protéine elle-même, qui diffuse dans la cellule depuis
son site de synthèse jusqu’à son site de liaison sur l’ADN.
*
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
1
/
47
100%