Respiration

Respiration
1 : Anatomie et histologie de l'appareil respiratoire
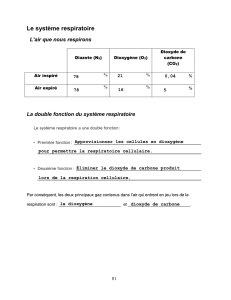
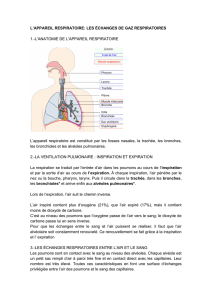
La fonction respiratoire consiste à capter l’oxygène atmosphérique dans le sang et à éliminer
le dioxyde de carbone et la vapeur d’eau. Les échanges se font à travers la barrière alvéollo-
capillaire et concernent des gaz.
Les alvéoles sont les unités fondamentales de l’appareil respiratoire mais les poumons n’ayant
pas de mobilité propre sont mobilisés par l’action des muscles du thorax qui sont
indispensables aux échanges.
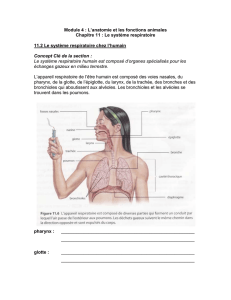
L’appareil respiratoire est formé de conduits de l’air (volume = 150 ml) et d’un tissu
fonctionnel impliqué dans les échanges : les petites bronchioles et les alvéoles.
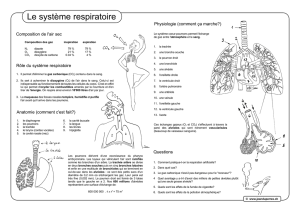
1. 1. Les voies conductrices
Les voies conductrices assurent le passage de l’air et non les échanges gazeux. Leur
volume est de 150 ml « espace mort ». Le rôle de ces conduits est de réchauffer,
d’hydrater l’air et de le dépoussiérer par l’épithélium cilié.
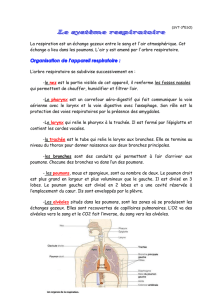
- les cavités nasales : réchauffement, humidification et filtration de l’air (les cils)
- le pharynx : carrefour aéro-digestif
- le larynx : rôle fondamental dans la phonation

- la trachée : 12 cm de longueur, 12 mm de diamètre, formée de 16 à 20 anneaux
cartilagineux. La trachée est tapissée de muqueuse faite d’un épithélium cylindrique à deux
couches dont la dernière est faite de cellules ciliées avec des cellules à mucus. Les fibres
élastiques permettent une certaine flexibilité et un retour à la normale après dilatation des
bronches. Le chorion peut également renfermer un tissu lymphoïde appelé MALT (Tissu
Lymphoïde Associé aux Muqueuses) ; il se compose de lymphocytes dispersés ou regroupés
en follicules lymphoïdes primaires ou secondaires. La trachée se divise en deux bronches
souches desservant chacune un poumon.
Composition et histologie de la trachée
- les bronches : Les bronches sont accompagnées de veines et artères, l’ensemble formant
le pédicule pulmonaire. Au niveau du hile, chaque bronche souche se divise en autant de
bronches lobaires qu’il y a précisément de lobes (3 à droite et 2 à gauche) ; la structure des
bronches est la même que celle de la trachée, sauf que les anneaux cartilagineux sont
complets.
Ensuite les bronches se subdivisent en éléments de plus en plus petits jusqu’à
l’élément le plus petit appelé bronchiole. On décrit la bronchiole terminale au centre
des lobules à mesure que le diamètre des bronches diminue, le cartilage se fragmente
puis disparaît. Il ne reste que du tissu fibro-musculaire. Les bronchioles entrent dans
un lobule, tissu fonctionnel.

Structure des voies de conduction
1.2. Les poumons
Le poids moyen des poumons est d’environ 800 g. Ils sont divisés en 3 lobes à droite, deux
lobes à gauche, desservis par les bronches lobaires. Dans la cage thoracique, les poumons sont
enveloppés dans les plèvres (séreuse) avec un feuillet viscéral collé au tissu pulmonaire et un
feuillet pariétal uni à la paroi thoracique. Entre les deux feuillets, on note la présence de la
cavité pleurale, elle contient du liquide pleural permettant le glissement (mobilisation des
poumons).

1.2.1 Histologie du parenchyme pulmonaire
Le tissu fonctionnel est subdivisé en lobules par des cloisons fibro-élastiques contenant les
vaisseaux et les bronches. Les lobules sont nombreux à la périphérie du poumon, ils sont rares
au centre.
- Les lobules pulmonaires
Un lobule pulmonaire est centré autour d’une bronchiole terminale. Chaque bronchiole
terminale se subdivise en plusieurs bronchioles respiratoires longues de 0,06 à 0,2 mm de
diamètre et qui se terminent par des sacs alvéolaires A une bronchiole terminale font suite 200
alvéoles (échanges gazeux). Les structures issues de la bronchiole terminale forment l’acinus.
Un acinus zone d’échange gazeux (ou lobule pulmonaire primaire), il est constitué d’une
bronchiole respiratoire donnant naissance à 3 à 6 canaux alvéolaires. Chaque canal alvéolaire
lui même se divise 2 à 3 fois. Chaque canal alvéolaire final s’ouvre au niveau de 2 ou 3 sacs
alvéolaires. A ce niveau le muscle disparaît, l’armature est uniquement faite de fibres
collagènes et élastiques. Les fibres élastiques permettent aux alvéoles de s’élargir lors de
l’inspiration. Les alvéoles se contractent ensuite "passivement" lors de l’expiration. Les fibres
de collagène évitent une trop grande distension des murs alvéolaires et donc leur rupture.

Les alvéoles sont des "sacs" de 200μm de diamètre. Leur structure histologique est adaptée à
leur fonction essentielle : l’échange oxygène/gaz carbonique. La surface d’échange de
l’ensemble des 300 millions d’alvéoles représente à peu près 140 m2 ; 12000 L d'air filtrés
face à 6000 L de sang perfusés par jour. Dans chaque alvéole on distingue la lumière
alvéolaire bordée par un épithélium alvéolaire et le "mur" ou cloison inter alvéolaire
(interstitium) où circule un réseau capillaire abondant.
Membrane alvéolo-capillaire
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
1
/
20
100%