THÈSE

N° d’ordre : 2941
- 2004 -
THÈSE
présentée à
L'UNIVERSITÉ BORDEAUX I
École Doctorale des Sciences de la Vie, Géosciences et Sciences de
L’Environnement
par
Carole MONTERRAT
Pour obtenir le grade de
DOCTEUR
SPÉCIALITÉ : SCIENCES DES ALIMENTS ET NUTRITION
*********************
LES SYNAPTOTAGMINES ET LE TRANSPORT VÉSICULAIRE
DANS LES CELLULES β
ββ
β PANCRÉATIQUES :
UNE ISOFORME POUR CHAQUE ÉTAPE ?
*********************
Soutenue le 10 Décembre 2004
Après avis de Messieurs :
Dominique BATAILLE, Directeur de Recherche INSERM U376, Montpellier
Michael SEAGAR, Directeur de Recherche INSERM UMR641, Marseille
Devant la commission d'examen formée de Messieurs:
Paul HIGUERET, Professeur Université Bordeaux 1
Dominique BATAILLE, Directeur de Recherche INSERM U376, Montpellier
Michael SEAGAR, Directeur de Recherche INSERM UMR641, Marseille
Marc LANDRY, Professeur Université Bordeaux 2
Jochen LANG, Professeur Université Bordeaux 1


Ce travail a été réalisé au laboratoire de Dynamique Moléculaire et Protéomique
fonctionnelle des Cellules Sécrétoires, au sein de l’Institut Européen de Chimie et
Biologie de Bordeaux.
Je voudrais remercier Jochen Lang, le courageux directeur qui a pris le risque de
prendre une “vieille” thésarde, grâce à lui j’ai pu réaliser cette thèse.
Je remercie messieurs D. Bataille et M. Seagar d’avoir accepté de juger mon travail et
messieurs P.Higueret et M.Landry d’avoir fait partie de mon jury.
Ce travail n’aurait pas été réalisable sans l’aide de Marie Noëlle Benassy et
Alexandra Milochau, je les remercie beaucoup pour l’aide précieuse qu’elles m’ont
apportée.
Pendant ces quatre années, j’ai bien entendu noué des relations de travail
enrichissantes, elles se sont doublées de relations amicales.
Je suis ravie de retrouver Sophie dans mes nouvelles fonctions, en espérant que
nous ayons autant de fous rires que lorsque nous avons nettoyé les étagères des
placards… Je pense que nous allons avoir du pain sur la planche, ou du beurre sur la
tranche, peut être du beurre sur la planche ???
Je pense que le flambeau du thésard-Conchita/Conchito a été maginifiquement
repris par Fred et Florence. Bravo ! Nous pouvons partir tranquilles. Florence est aussi
devenue madame Synaptotagmine 9, je te souhaite beaucoup de bonheur avec cette
merveilleuse protéine. Merci à Fred qui m’a bien aidée au moment où le sujet en avait
besoin mais surtout pour les franches rigolades (du pain sur la planche, ou du beurre
sur la tranche, peut être du beurre sur la planche ???).
Bien sur il y a aussi Valérie qui a toujours su me remonter le moral quand il le
fallait, tes conseils toujours justes m’ont fait avancer, merci beaucoup. Je reviendrai
sans doute te demander encore et encore et encore…

Merci aussi à Mathilde, incroyablement constante dans sa bonne humeur. Comment
fais-tu pour manger un clown tous les matins ? Il faudra que tu me donnes ton
fournisseur.
Bien entendu il y a eu Céline et Béatrice. Comment vous dire merci en deux
phrases ? Je crois simplement que l’année 2004 n’aurait pas été vivable sans vous. Je
sais que vous avez vécu tout ça avec moi, vous devriez avoir votre nom sur la première
page…
Merci à Philippe pour son soutien infaillible, à n’importe quelle heure du jour et de
la nuit. Vraiment la meilleure hot-line que je connaisse (quand il n’est pas sorti manger
du pain) !
A mes parents qui ont toujours soutenu mes choix, études-travail-études. Combien
de fois vous ai-je dit que c’était la dernière année ? C’est promis cette fois-ci c’est la
dernière...je crois !

Sommaire
- 1 -
INTRODUCTION ............................................................................................................11
I / LES MÉCANISMES CELLULAIRES DE LA SÉCRÉTION D’INSULINE ...............12
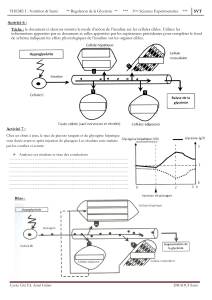
A / L’homéostasie du glucose ______________________________________________________ 12
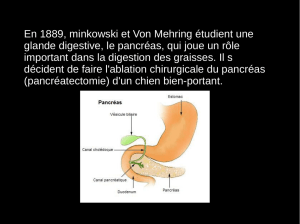
B / Le pancréas _________________________________________________________________ 14
C / De l’augmentation de la glycémie à la sécrétion d’insuline_____________________________ 16
1 – Le métabolisme du glucose dans les cellules β.........................................................................16
2 – La cellule β : une cellule spécialisée .........................................................................................17
D / Les systèmes d’amplification de la sécrétion d’insuline _______________________________ 18
1 – Les incrétines ............................................................................................................................18
2 – L’acétylcholine..........................................................................................................................20
3 – Les régulations autocrines et paracrines de la sécrétion d’insuline...........................................21
4 – Les sulfonylurées ......................................................................................................................22
E / La sécrétion pancréatique d’insuline ______________________________________________ 22
1 – Le synchronisme au sein d’un îlot.............................................................................................22
2 – La sécrétion pancréatique d’insuline, mesurée en fonction du temps, est biphasique...............24
3 – Les facteurs de régulation de la sécrétion indépendante des canaux K
+
dépendants de l’ATP .28
F / Le diabète de type 2 ___________________________________________________________ 29
II / LES MÉCANISMES MOLÉCULAIRES DE L’EXOCYTOSE...................................31
A / La biosynthèse de l’insuline_____________________________________________________ 31
1 – Transcription du gène et traduction de la préproinsuline ..........................................................31
2 – Maturation de la protéine et formation des vésicules sécrétoires ..............................................31
B / Les vésicules sécrétoires _______________________________________________________ 35
1 – Les différents types de vésicules dans les cellules β.................................................................35
1.1 – Les microvésicules de type synaptiques ou MVTS (Synaptic Like MicroVesicles) .........35
1.2 – Les granules sécrétoires ou vésicules larges à cœur dense (VLCD)..................................35
2 – Les étapes vers la fusion............................................................................................................36
2.1 – “Docking” ou amarrage.....................................................................................................36
2.2 – “Tethering” ou accrochage ................................................................................................38
2.3 – “Priming” ..........................................................................................................................38
3 – La fusion de la vésicule avec la membrane plasmique..............................................................39
3.1 – Fusion complète ou incomplète (“kiss-and-run”)..............................................................39
3.2 – Le pore de fusion : lipidique ou protéique ?......................................................................40
C / Les acteurs moléculaires de la fusion membranaire___________________________________ 41
1 – Le complexe SNARE ................................................................................................................41
1.1 – Mise en évidence du complexe de fusion universel ..........................................................41
1.2 – Description du complexe SNARE.....................................................................................43
1.3 – Les protéines SNARE et les cellules β pancréatiques .......................................................43
1.4 – Les protéines SNARE et la fusion membranaire ...............................................................45
2 – Les régulateurs de l’association du complexe SNARE .............................................................47
2.1 – La complexine ...................................................................................................................48
2.2 – Munc-18 ............................................................................................................................48
2.3 – Les protéines Rab ..............................................................................................................48
3 – Le calcium : facteur déclencheur de la stimulation ...................................................................50
4 – Les senseurs calciques...............................................................................................................52
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
1
/
216
100%