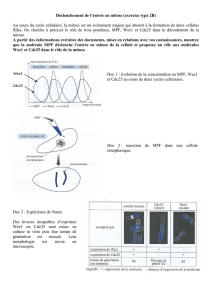
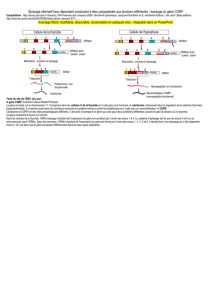
expression des différents variants dans des

AVERTISSEMENT
Ce document est le fruit d'un long travail approuvé par le jury de
soutenance et mis à disposition de l'ensemble de la
communauté universitaire élargie.
Il est soumis à la propriété intellectuelle de l'auteur. Ceci
implique une obligation de citation et de référencement lors de
l’utilisation de ce document.
D'autre part, toute contrefaçon, plagiat, reproduction illicite
encourt une poursuite pénale.
Contact : ddoc-theses-contact@univ-lorraine.fr
LIENS
Code de la Propriété Intellectuelle. articles L 122. 4
Code de la Propriété Intellectuelle. articles L 335.2- L 335.10
http://www.cfcopies.com/V2/leg/leg_droi.php
http://www.culture.gouv.fr/culture/infos-pratiques/droits/protection.htm

UNIVERSITÉ PAUL VERLAINE – METZ
UFR SCIENCES FONDAMENTALES ET APPLIQUÉES
ÉCOLE DOCTORALE « BIOLOGIE, SANTÉ, ENVIRONNEMENT »
THÈSE
Présentée et soutenue publiquement pour l’obtention du titre de
DOCTEUR DE L’UNIVERSITÉ PAUL VERLAINE – METZ
Mention : « Sciences de la Vie et de la Santé »
par
Hélène ALBERT
Le 26 octobre 2011
MEMBRES DU JURY
Rapporteurs
Professeur Bernard DUCOMMUN, Université Paul Sabatier, Toulouse
Professeur Mathias MONTENARH, Université de la Sarre, Hombourg, Allemagne
Examinateurs
Professeur Denyse BAGREL, Université Paul Verlaine, Metz (Directrice de thèse)
Docteur Eric BATTAGLIA, Université Paul Verlaine, Metz (Co-directeur de thèse)
Membre invité
Professeur Carolino MONTEIRO, Université de Lisbonne, Portugal
Laboratoire d’Ingénierie Moléculaire et Biochimie Pharmacologique (LIMBP)
Université Paul Verlaine - Metz, EA3940, Directeur Pr G. Kirsch
UFR SciFA, Campus Bridoux, Rue du Général Delestraint, 57070 METZ
Expression des différents variants dans des modèles
cellulaires et tissulaires de cancer mammaire
et régulation en conditions de stress génotoxique
ÉPISSAGE ALTERNATIF DES PHOSPHATASES CDC25
DANS LE CANCER DU SEIN :


Ce travail de doctorat
a été réalisé
au Laboratoire d’Ingénierie Moléculaire et Biochimie Pharmacologique
dans l’équipe de Biochimie Pharmacologique
UFR Sciences Fondamentales et Appliquées
Université Paul Verlaine - Metz
sous la direction du Pr Denyse BAGREL et l’encadrement du Dr Eric BATTAGLIA
a fait l’objet
d’une collaboration qui a nécessité un séjour de 5 semaines
au Laboratoire de Génétique Humaine de la Faculté de Pharmacie
et à l’Ecole Supérieure de Technologie de la Santé
Université de Lisbonne Ŕ Portugal
sous l’encadrement du Pr Carolino MONTEIRO et du Pr Miguel BRITO
Cette étude a été subventionnée
par la Ligue contre le Cancer (comité 57)
et le Conseil Régional de Lorraine
J’ai également bénéficié à titre personnel
d’une allocation de recherche attribuée par
le Ministère de l'Enseignement Supérieur et de la Recherche

Remerciements
REMERCIEMENTS
À Madame le Professeur Denyse Bagrel, pour m’avoir accueillie dans son équipe, pour
m’avoir guidée dès mes stages pré-doctoraux et pour m’avoir encouragée à me lancer dans
cette aventure. Je souhaite lui témoigner toute ma gratitude pour son soutien, sa rigueur
scientifique et sa grande bienveillance à mon égard. Je garderai de vous le souvenir d’une
femme courageuse et humaine.
Un grand merci au Docteur Eric Battaglia pour ses grandes qualités scientifiques et surtout
humaines. Je te suis très reconnaissante d’avoir su me remonter le moral dans les moments de
doutes par ton humour, ta confiance en moi, ton optimisme et tes intarissables nouvelles
idées.
Je suis très sensible à l’honneur que me font Messieurs les Professeurs Bernard Ducommun et
Mathias Montenarh d’avoir accepté d’être rapporteurs de cette thèse. Leur expertise dans le
domaine du cycle cellulaire ainsi que leur jugement apporteront une contribution précieuse à
ce travail.
À Monsieur le Professeur Carolino Monteiro grâce à qui nous avons pu acquérir les
échantillons tissulaires indispensables à notre étude et pour ses nombreux conseils. Je tiens
également à le remercier pour son accueil chaleureux lors de mon séjour dans son laboratoire
à Lisbonne, ainsi que toute son équipe en particulier Susana Santos pour son aide précieuse en
bioinformatique, ses conseils pratiques et sa gentillesse.
Merci également à Monsieur le Professeur Miguel Brito pour m’avoir accueillie dans son
laboratoire afin de m’initier à la pratique de la PCR en temps réel et pour nos discussions
enrichissantes. Je garderai un très bon souvenir de ce séjour grâce à son amabilité et à celle
des membres de son équipe.
Je tiens à remercier Monsieur le Docteur Marc Diederich, directeur du Laboratoire de
Biologie Moléculaire et Cellulaire du Cancer à Luxembourg, pour m’avoir permis d’utiliser
leur matériel de PCR en temps réel. Merci également à Sébastien Châteauvieux pour ses
conseils pratiques et à toute l’équipe du LBMCC pour leur sympathie à mon égard.
Je remercie aussi Madame le Professeur Pascale Bauda, directrice du Laboratoire des
Interactions Ecotoxicologie, Biodiversité, Ecosystèmes, de nous avoir permis d’avoir accès à
leur équipement de PCR en temps réel. Merci également à Yann pour sa sympathie.
À Monsieur le Professeur Armand Maul, biostatisticien à l’Université Paul Verlaine-Metz,
pour sa disponibilité et son aide précieuse dans le choix des tests statistiques les plus adaptés à
notre étude clinique.
À nos collègues de l’équipe d’Ingénierie Moléculaire pour leur convivialité et leur
approvisionnement en éthanol, en azote liquide et en divers produits chimiques.
À tous mes collègues de l’équipe de Biochimie Pharmacologique passés et présents :
doctorants, stagiaires, techniciennes, maîtres de conférence, qui ont contribué d’une façon ou
d’une autre à l’agréable ambiance qui a régné au cours de ces années passées au laboratoire.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
1
/
222
100%