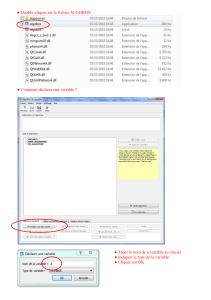
π A - WebCampus

L’INFÉRENCE PHYLOGÉNÉTIQUE
Raphaël Helaers , Ph.D.
Bioinformaticien ICP-UCL

PHYLOGÉNIE
« Étude de la formation et de l’évolution des
organismes vivants en vue d’établir leur parenté »

PLATON, ARISTOTE (ANTIQUITÉ)
•Classification des organismes vivants
•Échelle permettant de classer ces organismes
des plus simples aux plus perfectionnés
•Dans cette vision, les espèces sont fixes et
permanentes
•Incompatible avec la notion d’évolution

LINNÉ (1758) : TAXONOMIE
Règne - Animal
Phylum - Chordata
Subphylum - Vertebrata
Classe - Mammalia
Ordre - Carnivora
Famille - Ursidae
Genre - Ursus
Espèce - U. maritimus

DARWIN (1859)
•« De l’Origine des Espèces »
•Hérédité : les espèces partagent un lien de
parenté
•La diversité du vivant s’explique par le
processus d’évolution
•Moteur de l’évolution adaptative : la sélection
naturelle
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
1
/
65
100%