III) La 3ème semaine : Du 15 ème jour au 21ème

III) La 3 ème
semaine : Du 15 ème
jour au 21 ème
jour inclus
Durant cette semaine, on a la réalisation d’un embryon discoïde, planiforme et tridermique, la mise en
place d’un axe de symétrie céphalo-caudal avec l’apparition de la chorde et des modifications spatiales du disque
avec les prémices de la délimitation de l’embryon.
A- La gastrulation : mise en place de l’endoblaste et du mésoblaste intra-embryonnaire
B- La formation de la chorde et les modifications de mésoblaste intra embryonnaire
C- L’apparition de l’allantoïde, la mise en place des cellules germinales primordiales, des
ébauches vasculaires extra et intra-embryonnaires et du SNC (La neurulation primaire)
D- La mise en place du SNC : neurulation primaire
A- La gastrulation
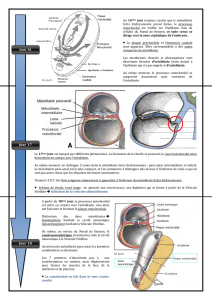
Au 15ème jour, du développement, les cellules épiblastiques de surface se multiplient abondamment
pour former une saillie qui se développe dans un plan médian et prend naissance au niveau de la région caudale.
Cette saillie progresse vers la région céphalique (sans l’atteindre) et est creusée d’une gouttière à son centre; il
s’agit de la ligne primitive. La ligne primitive comprend, dans sa région céphalique, une zone épiblastique qui
s’est multipliée et qui concourt à former une masse volumineuse, un bourrelet, c'est le nœud de Hansen, qui
apparaît au 16ème jour du développement et au centre duquel se trouve le prolongement de la gouttière creusée
dans la ligne primitive. (→ Fig. 3.1)
Au niveau des berges de la gouttière, des cellules épiblastiques qui se sont multipliées se détachent, et se
transforment du point de vue morphologique : de cylindriques elles deviennent étoilées, avec des limites
cellulaires floues, un noyau ovalaire, nucléolé et une chromatine fine. Ces cellules épiblastiques modifiées dans
leur structure s’invaginent entre l’hypoblaste et l’épiblaste, ainsi elles vont migrer pour s’interposer
progressivement entre l’épiblaste et l’hypoblaste ; elles cheminent entre les deux feuillets pour les séparer. (→ Fig.
3.2) Une fois qu’elles se sont invaginées, elles élaborent des constituants macromoléculaires d’une substance
fondamentale dont elles s'entourent, et donc de proche en proche entre l’hypoblaste et l’épiblaste se met en
place progressivement un tissu constitué d’une substance fondamentale très abondante, d’aspect gélatineux, au
sein de laquelle sont dispersées quelques cellules : le mésoblaste intra-embryonnaire, qui a une structure
identique à celle du mésoblaste extra-embryonnaire mais dont la localisation anatomique diffère.
Pendant que cela s’effectue, au fond de la gouttière se trouvent des cellules épiblastiques modifiées qui
vont venir au contact des cellules hypoblastiques et s'insinuer entre elles après les avoir décollées (→ Fig. 3.2).
Dès lors, ces cellules changent de morphologie, elles deviennent cubiques, prennent une organisation épithéliale
et se substituent petit à petit à l’hypoblaste. Lorsque toutes les cellules hypoblastiques seront remplacées, c'est-à-
dire sur tout le disque et la paroi de la vésicule vitelline secondaire, l’hypoblaste n’existe plus : on parle
d’entoblaste, qui devient le 2ème feuillet de l’embryon.
Une fois que le mésoblaste intra-embryonnaire et l'entoblaste sont mis en place, l’épiblaste prend le nom
d’ectoblaste; on obtient alors un embryon discoïde, planiforme et tridermique comportant : l’ectoblaste en
rapport avec la cavité amniotique, le mésoblaste intra-embryonnaire, et l’entoblaste en rapport avec la cavité
de la vésicule vitelline secondaire (→ Fig. 3.3)
Le mésoblaste intra-embryonnaire, lors de sa migration à partir des cellules filles s’insinue partout entre
l’ectoblaste et l’endoblaste sauf en 2 zones : la membrane pharyngienne dans la région céphalique, et la
membrane cloacale dans la région caudale. Au niveau de ces 2 membranes, il n’y a jamais d’interposition de
mésoblaste intra-embryonnaire entre l’ectoblaste et l’endoblaste, car ce sont des membranes didermiques au
niveau desquelles l’ectoblaste et l’endoblaste sont soudés et donc où le mésoblaste intra-embryonnaire ne peut
pas migrer. Ainsi le mésoblaste intra-embryonnaire s’insinue partout en avant et en arrière des deux membranes.
(→ Fig. 3.1)
Il faut noter que l’ectoblaste est en contact avec l’amnios au niveau des jonctions amnio-
ectoblastiques ; latéralement, en périphérie du disque, les mésoblastes intra et extra-embryonnaire sont
raccordés : la somatopleure extra-embryonnaire en rapport avec l’amnios et la splanchnopleure extra-
embryonnaire avec la paroi de la vésicule vitelline secondaire (→ Fig. 3.3).
Durant la troisième semaine, lorsque le mésoblaste intra-embryonnaire devient autonome c'est-à-dire
que ses cellules se multiplient par mitose, la ligne primitive régresse et disparaît presque totalement : à la fin de
celle-ci, elle n’est plus que vestigiale. Toutefois, avant sa disparition complète dans le courant de la 4ème semaine,
la ligne primitive donne naissance, dans la région caudale de l’embryon qui est en train de se développer, dans
une zone médiane et para-médiane, à une masse volumineuse constituée des cellules mésoblastiques tassées

formant une structure en corde appelée l’éminence ou le bourgeon caudal. Ainsi dans des conditions normales,
la ligne primitive disparaît, mais dans certains cas elle persiste et donne lieu à des malformations, des tumeurs
situées dans la région coccygienne avec un diagnostic très redoutable, les tératomes sacro-coccygiens.
B- La formation de la chorde
La chorde est très importante, car elle va former l’axe de symétrie céphalo-caudal du futur embryon.
Sa formation s’effectue en trois stades : • 16ème jour : canal chordal
• 17ème jour : plaque chordale
• 18ème jour : chorde pleine
Au 16ème jour, au niveau de l'orifice du nœud de Hansen, les cellules épiblastiques qui s'étaient développées
à partir de la ligne primitive se regroupent et s’invaginent en doigts de gant dans un plan médian, en direction de
la région céphalique, pour former un canal. Les cellules se disposent de manière épithéliale, deviennent cubiques
et forment la paroi du canal chordal, aussi appelé prolongement céphalique de la ligne primitive, car il
progresse du nœud de Hansen vers la région céphalique : l'orifice se situe au niveau du nœud de Hansen et
l'extrémité borgne à l'opposé, écarte l’ectoblaste et l’entoblaste en progressant vers la membrane pharyngienne
(→ Fig. 3.4).
Le mésoblaste intra-embryonnaire a pu migrer en avant de la membrane pharyngienne et en arrière de la
membrane cloacale et du canal chordal. (→ Fig. 3.4) L’extrémité céphalique (borgne) du canal comporte une
partie mésoblastique très dense qui forme une saillie où les éléments se densifient, formant la plaque pré-
chordale qui se dirige vers la membrane pharyngienne. De fait, le prolongement céphalique de la ligne primitive
qui se trouve entre les 2 feuillets est formé par le canal chordal, à l’extrémité duquel se trouve la plaque pré-
chordale, l’ensemble progressant vers la membrane pharyngienne. Sur une coupe transversale, on remarque que
le plafond du canal chordal est en rapport avec l’ectoblaste, son plancher avec l’entoblaste et ses parois latérales
avec le mésoblaste intra-embryonnaire. (→ Fig. 3.5)
Au 17ème jour, la plaque pré-chordale vient buter contre la membrane pharyngienne qui est un obstacle
infranchissable : la progression du canal chordal s’arrête, car dans la membrane pharyngienne l’endoblaste et
l’ectoblaste sont soudés. De plus, à cause de la pression exercée par le canal, le plancher du canal et l’entoblaste
sous-jacent dégénèrent de la région céphalique à la région caudale jusqu’à disparaître complètement, ne
persistent alors que le toit du canal chordal et ses parois latérales : on est alors au stade de plaque chordale. (→
Fig. 3.6) Il demeure toutefois un lieu de communication entre la cavité amniotique et la vésicule vitelline
secondaire : le canal neurentérique. Les rapports de la plaque chordale sont les suivants : son toit est en
rapport avec l'ectoblaste, ses parois latérales avec le mésoblaste intra-embryonnaire et ses bords se rattachent à
l'entoblaste. Dans le plan médian, cette plaque forme le toit de la vésicule vitelline secondaire (→ Fig. 3.6).
Au 18ème jour, la chorde se forme; elle résulte d’une transformation de la plaque chordale en cordon
chordal plein. D'une part, on observe un mouvement de striction des bords de la plaque, vers l'intérieur,
tellement puissant que les bords de la plaque se détachent de l’entoblaste. D'autre part, on constate de
nombreuses mitoses au niveau des parois de la plaque donc les cellules s’accumulent ce qui entraîne donc
l’épaississement des parois. Par ailleurs, le phénomène de striction devient de plus en plus important et les
extrémités, où les cellules prolifèrent, vont alors se rapprocher jusqu’à fusionner l’une avec l’autre, formant une
cavité temporaire, puis à terme, les bords de la plaque se soudent et toutes les cellules remplissent complètement
le cordon qui vient de se former, donc la cavité disparaît. Enfin, au niveau de l’entoblaste se déroulent aussi des
proliférations cellulaires; les cellules vont à la rencontre les unes des autres pour former une zone d’entoblaste
régénéré. (→ Fig. 3.7)
On a alors un cordon cellulaire dénué de toute cavité, dans l’axe médian de l’embryon : la chorde, qui
s’étend du nœud de Hansen (en train de disparaître) à la membrane pharyngienne, formant l’axe de symétrie
céphalo-caudal de l’embryon. La paroi dorsale de la chorde est en rapport avec l’ectoblaste, sa paroi ventrale
avec l’endoblaste et ses parois latérales avec le mésoblaste intra-embryonnaire (→ Fig. 3.8).
Entre le 18ème et le 21ème jour, le disque embryonnaire se développe de manière asymétrique : il croît
beaucoup plus au niveau céphalique qu’au niveau caudal. La ligne primitive et le nœud de Hansen régressent : au
21ème jour, la distance entre le nœud de Hansen et la membrane cloacale est minime par rapport à la distance
entre la membrane cloacale et la membrane pharyngienne ; on peut admettre que la chorde s'étend de la
membrane cloacale à la membrane pharyngienne, dans un plan médian. (→ Fig. 3.9). D’autres

modifications vont concerner le mésoblaste intra-embryonnaire : dans certaines zones, les cellules se multiplient
abondamment, en particulier au voisinage de la chorde. En fonction de l'importance de la prolifération, on
distingue trois massifs cellulaires, qui suivent un gradient de volume décroissant de la chorde vers la périphérie
du disque (→ Fig. 3.10) :
le mésoblaste para-axial, abondant et compact, proche de la chorde (axe de référence), là où les cellules se
sont le plus multipliées
le mésoblaste intermédiaire, deuxième massif, moins volumineux, car les cellules mésoblastiques se sont
moins multipliées, situé entre le mésoblaste para-axial et le 3ème massif
le mésoblaste latéral, le moins volumineux et le plus éloigné de l’axe, en rapport avec le mésoblaste extra-
embryonnaire qui revêt respectivement l’amnios (somatopleure) et la paroi de la vésicule vitelline secondaire
(splanchnopleure).
Le mésoblaste para-axial s'étend donc de la région céphalique à la région caudale et encadre la chorde ;
à la fin de la 3ème semaine, vers le 20ème jour, il se segmente dans sa région céphalique en petits amas d’égal
volume, les métamères, qui se disposent en piles d’assiettes les uns au-dessous des autres. Ce processus de
division, la métamérisation, donne ainsi naissance aux quatre premières paires de somites, les somites
occipitaux. Au cours de la quatrième semaine, la métamérisation gagne l’ensemble du mésoblaste para-axial, et
les paires de somites s'ajoutent les unes aux autres.
C- La mise en place de l’allantoïde
L’allantoïde se forme au 18ème jour du développement, dans la partie caudale de l’embryon, et dans un
plan médian : il s'agit d'une évagination émise par l'entoblaste au niveau de la paroi de la vésicule vitelline
secondaire. Son extrémité est borgne, il comporte une lumière et une paroi formée d'entoblaste, tapissée de
mésoblaste extra-embryonnaire (→ Fig. 3.11).
L'allantoïde revêt plusieurs intérêts :
les ébauches vasculaires vont se former dans le mésoblaste extra-embryonnaire qui est en rapport avec
l’allantoïde
lors de la 4è m e semaine du développement, lors de la délimitation, l’allantoïde fait partie du pédicule de
fixation et va être incorporé dans le cordon ombilical
l’allantoïde dégénère et donne naissance à une partie longitudinale de la vessie, le trigone vésical.
D- La mise en place des ébauches vasculaires extra et intra-embryonnaires
Ce phénomène est très important : si cette mise en place est anormale, le développement n’est plus
possible. Au 18ème jour, dans le mésoblaste extra-embryonnaire, en rapport avec la paroi de la vésicule vitelline
secondaire, et dans le mésoblaste intra-embryonnaire, en rapport avec l’entoblaste qui revêt l’allantoïde, se
forment des vaisseaux.
Le mésoblaste extra-embryonnaire est constitué de cellules très espacées, mais dans certains endroits,
celles-ci se regroupent et forment des amas, ou îlots, constitués d’éléments mésoblastiques tassés les uns contre
les autres et d’une substance fondamentale peu développée. Au sein de ces îlots apparaissent de petites cavités
cellulaires qui confluent pour former une cavité vasculaire, unique. La paroi de cette cavité est formée des
cellules mésoblastiques de la périphérie des îlots, modifiées, très aplaties, qui sont unies les unes aux autres et
forment un épithélium, et qu'on appelle angioblastes. Ce phénomène correspond donc à la formation d'un
vaisseau. En même temps, les cellules situées au centre des îlots changent de morphologie ; elles deviennent
arrondies, à gros noyau, et se retrouvent dans la cavité vasculaire : ce sont les hémocytoblastes primordiaux,
qui possèdent une potentialité hématopoïétique, c'est-à-dire qu'elles sont capable de fabriquer des éléments
figurés du sang ou du moins leurs précurseurs (→ Fig. 3.12).
Les vaisseaux de faible calibre font fusionner entre eux, donnant des vaisseaux de plus fort calibre pour
former le premier système vasculaire extra-embryonnaire, le système vitellin. Il comporte des paires de
vaisseaux : 2 artères vitellines, 2 veines vitellines, et les ramifications capillaires qui en dépendent. Dans cette
région se trouvent les îlots de Wolff et Pander, qui sont certes angioformateurs, car ils forment des
vaisseaux, mais aussi sanguinoformateurs, puisqu’ils forment des éléments sanguins souches. Par conséquent,
le système vasculaire extra-embryonnaire vitellin a essentiellement un rôle hématopoïétique.
Pendant ce temps, dans le mésoblaste extra-embryonnaire qui entoure la paroi de l’allantoïde, les
éléments mésoblastiques se regroupent en îlots, qui se creusent de la même façon pour former une cavité qui se
développe pour former un deuxième système vasculaire extra-embryonnaire, le système chorio-allantoïdien,
qui est également pair : il comporte 2 artères chorio-allantoïdiennes, 2 veines chorio-allantoïdiennes et les

réseaux capillaires qui en dépendent. Il existe toutefois une grande différence entre ces deux systèmes, puisque
dans la région allantoïdienne ne se forment que des vaisseaux : le système chorio-allantoïdien est uniquement
angioformateur, on n'y observe jamais de formation d’éléments sanguins souches.
Toujours dans le même temps, dans le mésoblaste intra-embryonnaire se forment des îlots qui se
creusent pour former des cavités et donner naissance à des vaisseaux, formant le système vasculaire intra-
embryonnaire qui est aussi uniquement angioformateur. Dans l’embryon, ces vaisseaux se disposent d’une
manière particulière, et en particulier dans la zone céphalique. Dans le mésoblaste intra-embryonnaire situé de
part et d’autre de la membrane pharyngienne, là où le mésoblaste a pu migrer, se forment des îlots qui se
creusent sous forme de tubes, donnant les deux tubes cardiaques (un à droite et un à gauche), ainsi que les
vaisseaux artériels qui en partent et les vaisseaux veineux qui y arrivent. Chaque tube cardiaque possède deux
extrémités :
une extrémité artérielle d’où partent deux artères, les aortes, respectivement ventrale et dorsale
une extrémité veineuse où arrivent des veines, les veines cardinales antérieure et postérieure, provenant
respectivement de la région céphalique et de la région caudale de l'embryon.
Pour résumer, on trouve dans l’embryon : 2 tubes cardiaques, 2 aortes ventrales, 2 aortes dorsales et 4 veines
cardinales (2 antérieures et 2 postérieures).
Au 21ème jour du développement, les tubes cardiaques commencent à battre, il y a donc apparition d’un
mouvement circulatoire. De plus, les réseaux vasculaires extra-embryonnaires, respectivement vitellin et
chorio-allantoïdien, se connectent au réseau vasculaire intra-embryonnaire et donc les échanges (uniquement de
gaz à ce stade) entre le placenta maternel et le disque embryonnaire en développement sont possibles (→ Fig.
3.12)
E- La mise en place des cellules germinales primordiales
Entre le 18ème et le 21ème jour, au sein du mésoblaste extra-embryonnaire en rapport avec la zone où
l’allantoïde s'abouche à la vésicule vitelline secondaire, on voit apparaître des cellules globuleuses, très
volumineuses, avec un gros noyau sphérique, nucléolé, à chromatine assez fine en réseau et un cytoplasme riche
en phosphatases alcalines. Elles sont d’origine épiblastique, et s'étant modifiées dans leur forme et dans leur
métabolisme, elles forment les gonocytes primordiaux, dans le mésoblaste extra-embryonnaire qui entoure la
zone de l’allantoïde. Ce sont les cellules sexuelles primitives, qui naissent en dehors du disque embryonnaire.
Progressivement, durant la 4ème semaine, ces cellules migrent puis pénètrent dans l’embryon en formation,
jusqu'à atteindre au cours de la 5ème semaine la crête génitale, et s'y associant, formant les gonades.
F- La mise en place du SNC : la neurulation primaire
L a neurulation comporte deux grands stades, très disproportionnés car la première phase est
beaucoup plus importante : d'abord la neurulation primaire du 18ème au 28ème jour, puis la neurulation
secondaire du 28ème au 30ème jour.
La neurulation primaire est progressive, à cheval sur 2 semaines; elle comporte 3 étapes :
• 18ème jour : formation de la plaque neurale
• 20ème jour : formation de la gouttière neurale
• 22ème jour : formation du tube neural
Au 18ème jour, en avant du nœud de Hansen qui est en train de disparaître progressivement, et en
direction de la région céphalique, l’ectoblaste s'épaissit de manière considérable pour former une plaque en
forme de semelle, qui présente une asymétrie : au niveau de cet épaississement superficiel, situé dans la région
médiane, et qui déborde latéralement dans les régions para-médianes, le développement est plus important dans
la région céphalique que dans la région caudale. (→ Fig. 3.8) C'est la plaque neurale, formée de cellules
ectoblastiques qui se sont multipliées et qui ont des potentialités différentes puisqu’elles sont destinées à former
des neurones. Cet ectoblaste modifié, constitutif de la plaque neurale, porte le nom de neurectoblaste, et
donnera naissance au SNC. Le reste de l’ectoblaste est appelé ectoblaste superficiel et donnera l’épiderme,
qui revêt la peau de l’embryon.
Aux 19ème et 20ème jours, la région centrale de la plaque se déprime alors que les bords se surélèvent,
formant une gouttière neurale, largement ouverte dans la cavité amniotique (→ Fig. 3.13).
A la fin de la 3ème semaine, l’embryon est discoïde, planiforme et tridermique (ectoblaste, mésoblaste
intra-embryonnaire et entoblaste). Il présente un axe de symétrie céphalo-caudal de la membrane pharyngienne à
la membrane cloacale, la chorde ; des réseaux vasculaires se mettent en place, que ce soit à l'intérieur ou en
dehors de l'embryon ; le mésoblaste intra-embryonnaire a commencé à se modifier, notamment avec l'apparition
des somites ; la neurulation a commencé (→ Fig. 4.3)
1
/
4
100%