CYTOGENETIQUE GENERALE Chapitre II La Méiose

CYTOGENETIQUE GENERALE
Chapitre II La Méiose
Introduction
La première division cellulaire chez les eucaryotes que nous venons de voir est
la mitose qui concerne les cellules somatiques et assure la naissance de cellules
identiques à la cellule mère lors de la multiplication asexuée (elle conserve donc
l'information génétique).
La deuxième division cellulaire chez les eucaryotes que nous allons voir est
la méiose qui aboutit à la production de cellules sexuelles ou gamètes pour
la reproduction sexuée.
Chez les animaux, la méiose est un processus se déroulant durant
la gamétogénèse (spermatogenèse ou ovogenèse), c'est-à-dire durant
l'élaboration des gamètes (les spermatozoïdes chez le mâle et les ovules chez la
femelle) chez les espèces dites diploïdes.
Chez les végétaux, la méiose produit des grains de spores dans les étamines.
Elle donne des cellules haploïdes (cellules contenant n chromosomes) à partir de
cellules diploïdes (cellule contenant 2n chromosomes)
Chez l'homme, une cellule normale contient 2n = 46 chromosomes (donc 23
paires) alors qu'un gamète contient n = 23 chromosomes au cours de deux
divisions). Chez les espèces haploïdes (comme la Sordaria macrospora), la
méiose intervient après la fécondation pour diviser la cellule-œuf (à 2n
chromosomes). Mais en plus de ce rôle de division, la méiose a un rôle
important dans le brassage génétique (mélange des gènes) et ce, grâce à deux
mécanismes de brassage : le brassage inter-chromosomique et le brassage intra-
chromosomique).
Ainsi, durant la méiose, la quantité d'ADN au sein de la cellule évolue au cours
du temps.
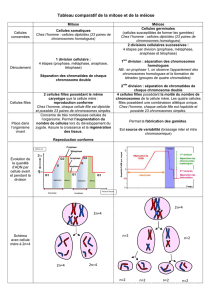
Chaque cellule va donc séparer son patrimoine génétique (contenu dans
des chromosomes) en deux afin de ne transmettre que la moitié de ses gènes aux
cellules filles.

Les différentes étapes de la méiose
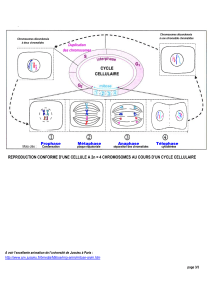
La méiose est un cycle cellulaire qui se déroule en plusieurs étapes regroupées
en deux divisions cellulaires, successives et inséparables. Les deux divisions
méiotiques sont :
- la division réductionnelle ou méiose I ou méiose réductionnelle.
- la division équationnelle ou méiose II ou méiose équationnelle.
A] La méiose I ou méiose réductionnelle
Pendant que dans la mitose nous partons d’une cellule 2n = 2C pour aboutir à
deux cellules filles 2n = 2C chacune, la méiose réductionnelle part d’une cellule
2n = 2C pour donner deux cellules filles 1n = 2C chacune.
Les phases de la méiose I sont :
- La prophase I
- La métaphase I
- L’anaphase I
- La télophase I
NB. La méiose est précédée d’une interphase, comme à la mitose, avec la phase
S où on a le dédoublement des chromosomes. Après cette interphase, les
différentes phases de la méiose I peuvent commencer.
1) La prophase I
C’est la phase la plus longue de la méiose I et aussi la plus longue de toute
la méiose. Elle se subdivise en 5 sous-phases : le leptotène, le zygotène, le
pachytène, le diplotène et la diacinèse.
Le Leptotène (du grec leptos = mince et taenia = le ruban)
Le leptotène marque le début de la condensation de la chromatine et
l’attachement des télomères (extrémités des chromosomes) à l'enveloppe
nucléaire par la plaque d'attache. Ici les chromosomes, dupliqués, sous
forme de filaments fins irréguliers deviennent visibles et individualisés
par la condensation. Cette condensation va continuer jusque à la fin de de
la prophase.
Ce début de condensation fait apparaître, le long de ces filaments fins, des
zones bien limitées en forme de spirale qui vont croissantes : les
chromomères. Ce sont des zones d’épaississement très colorables.

Au leptotène, le rapprochement de chromosomes homologues a
commencé mais aucune paire de chromosomes homologues n’est
observable.
Les centrioles, dédoublés, commencent leur migration vers les pôles.
Le zygoène (Zygos = joug et ten = filament)
C’est la seconde phase de la prophase I.
Au cours du zygotène,
La migration des centrioles aux pôles se poursuit.
La condensation se poursuit.
C’est le début de l’appariement des chromosomes homologues. C’est la
formation des synapsis.
NB dans une cellule diploïde, il y a un lot de chromosomes venant de la
mère et un lot venant de père. Un chromosome dans un lot a son
semblable dans l’autre lot. Les deux chromosomes ainsi identifiés sont
appelés des homologues.
L’appariement des chromosomes homologues se fait par la formation du
complexe synaptonémal ou synapton.
Il y a aussi une convergence des télomères vers le centromère (un peu
comme une fermeture éclair).
Les phénomènes de crossing-over (enjambement) s’observent dans cette
phase. A la fin, il y a organisation « en bouquet » des chromosomes.
L'ensemble des deux chromosomes homologues ainsi en bouquet prend le
nom de tétrade (car 4 chromatides réunies deux à deux par leur
centromères) ou un bivalent (car 2 chromosomes).
NB. Cette zygotène est caractéristique de la méiose. Elle n’existe pas dans
la mitose.
Le pachytène (Packhus = épais)
C’est la phase la plus longue de la prophase I (environ 2semaines pour des
spermatozoïdes humains).
Appariement strict des chromosomes homologues et apparition des
nodules de recombinaison et de nodules tardifs qui permettent les
enjambements ou crossing-over (échanges entre chromatides
homologues). Cette phase a une importance considérable dans le brassage
chromosomique.
Le stade pachytène est donc marqué par l'achèvement du synapsis c’est-à-
dire la formation des bivalents. Pour exemple : chez l’homme, les
chromosomes appariés constituent 22 bivalents autosomiques et
01bivalent sexuel condensé sous forme d'une vésicule sexuelle (VS) chez
le mâle. Les chromosomes se condensent progressivement jusqu'à

atteindre leur maximum ce qui permet d’observer les tétrades ou bivalents
qui sont plus courts et épais et nets.
Les échanges de matériel génétique entre chromosomes homologues est
bien visible. Les crossing-over sont bien visibles.
Le Diplotène(diploos = double)
Dans cette phase, les chromosomes, homologues, commencent à se
séparer avec la désintégration du complexe synaptonémal (désynapsis):
les centromères s'éloignent l'un de l'autre.
Les bivalents restent réunis (attachés) en certains points appelés chiasmas.
Au niveau de ces points deux des quatre chromatides semblent
s'entrecroiser (chiasma). Ces points sont généralement la manifestation
morphologique d'échange de matériel génétique entre deux chromatides
(de deux chromosomes homologues) c'est à dire le crossing-over.
Pour le bon déroulement de la méiose il en faut au minimum un crossing-
over par chromosome (en moyenne 2-3).
Il y a décondensation de la chromatine et formation des grandes boucles
permettant un fort taux de transcription.
Ce stade peut durer des jours ou même des années, selon le genre de
d'organisme et son type (ovocytes de 1er ordre dans l'ovaire bloqués dans
le stade diplotène jusqu'à leur transformation en ovocytes de 2e ordre,
c'est-à-dire de la vie fœtale à chaque cycle adule chez la femme). La
transcription des chromosomes est très active au cours du stade diplotène,
notamment chez les femelles car l'œuf se consacre activement au stockage
du matériel qui sera utilisé dans les toutes premières divisions du
développement embryonnaire (dans la segmentation).
La diacinèse (dia = à travers et kinesis = mouvement) :
C'est une étape de transition entre le stade diplotène et la métaphase.
Elle marque la fin de la prophase I.
Au cours de ce stade, les chromosomes se condensent à nouveau. Ici les
chromosomes homologues raccourcissent et se condensent une fois de
plus au maximum.
On observe la recondensation de la chromatine avec détachement des
télomères de l'enveloppe nucléaire et le glissement des chiasmas vers les
télomères (terminalisation des chiasmas). À la fin, il y a disparition de
l'enveloppe nucléaire.

Remarques
R1
Le diplotène de la prophase I peut durer plusieurs années chez l'ovocyte. En
effet au cours de ce stade, l'ovocyte I augmente de volume, la décondensation
des chromosomes permet la synthèse d'ARN messager et d'ARN
ribosomiques qui seront stockés dans le cytoplasme et serviront de réserve pour
le futurzygote lors des premières division de segmentation.
R2
Une des particularités de la méiose est la formation de bivalents.
Après cette longue phase qu’est la prophase I, on passe à la métaphase I
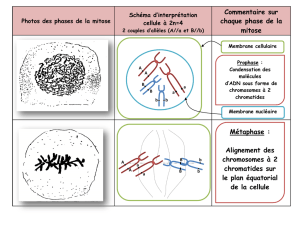
2) La métaphase I
La métaphase I, la membrane nucléaire et les nucléoles disparaissent. Les
paires de chromosomes homologues (bivalents) se placent de part et
d'autre du plan équatorial (PE). Pour chaque bivalent, les centromères se
placent de part et d'autre ainsi qu'à égale distance du plan équatorial. Leur
orientation se fait de façon aléatoire : on appelle ce phénomène la
« ségrégation indépendante ». Cette ségrégation permet un second degré
de diversification des cellules-filles : le brassage interchromosomique
 6
7
8
9
10
11
12
13
14
15
16
17
6
7
8
9
10
11
12
13
14
15
16
17
1
/
17
100%