Introduction

Physiologie repro. et dev. animal Mr Richard Semestre 5
1
Physiologie de la reproduction et du
développement animal
Introduction
La fonction de reproduction met en œuvre des
mécanismes à la fois extrêmement communs chez
tous les vertébrés, et à la fois différentiée par des
adaptations surprenantes, résultat de l’évolution
des espèces. Elles sont destinées à faire que
l’espèce se maintienne, dans des environnements
qui auront changé au cours du temps.
L’appartenance à un sexe d’un individu vertébré
est due à la présence de gènes spécifiques, qui
induisent la différenciation sexuelle. Chez
l’humain, la femme a XX, l’homme a XY.
On note la présence du gène SRY, responsable de la différenciation testiculaire. Le chromosome Y va
porter un nombre relativement restreint de ces gènes qui mènent à la différenciation.
Homme et femme ont l’X, et donc il doit porter des gènes codant pour des protéines qui doivent
servir aux deux. Chez la femelle on trouve de plus un deuxième X, alors on se demande ce qu’il va
apporter. En fait ce X « surnuméraire » ne sert pas à grand-chose, et la plupart du temps, il va se
retrouver sous forme de chromatine extra-condensée, inutile.

Physiologie repro. et dev. animal Mr Richard Semestre 5
2
On a dénommé cette chromatine corpuscule de Baar. Ca a été pendant longtemps un moyen de
déterminer le sexe d’un embryon, d’observer la présence du corpuscule de Baar. Il est possible de
trouver une femme portant 3 chromosomes X, c’est viable, et l’on observe alors 2 corpuscules de
Baar.
Les fonctions de reproduction sont assez similaires chez tous les vertébrés, et particulièrement chez
le mâle. On va trouver des différences structurales, aussi bien au niveau localisation
qu’anatomique, mais beaucoup de similitudes de fonctionnement.
La spermatogenèse est encore très conservée entre les espèces, que ce soit au niveau temporel ou
quantitatif.
Chez les femelles, on va trouver plus de différences, morphologiques et fonctionnelles.
L’ovogenèse, et son schéma de méiose, pour la constitution de l’ovocyte est similaire, suivant les
espèces. Le niveau temporel est par contre très différent suivant les espèces. Autour de l’ovocyte,
les réserves alimentaires sont très différentes. On aura parfois la constitution d’un vitellus, plus ou
moins présent, plus ou moins abondant. L’ovulation, en termes de quantité émise sera très variable.
1 chez l’homme, à une vingtaine chez les rongeurs. Derrière ça, si l’on a une quinzaine d’ovocytes
émis, et fécondés, il faudra pouvoir les accueillir, et donc il y aura des différenciations structurales,
morphologiques et fonctionnelles suivant les espèces. On a parfois des possibilités d’activation,
dans certaines espèces peu évoluées, par parthénogenèse, c'est-à-dire sans fécondation.
Il y aura différenciation de production des hormones.
Si l’on prend des espèces comme le lapin, le chat, il n’y aura pas d’ovulation tant qu’il n’y a pas de
fécondation (post-coïtal). Chez la chauve-souris, il y aura un décalage extraordinaire entre
fécondation et ovulation, jusqu’à 3 mois. La femelle peut stocker de la semence mâle jusqu’à ce que
les conditions soient suffisantes pour la gestation.
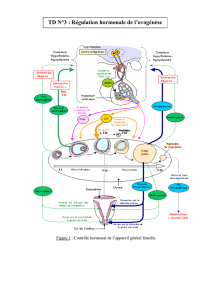
Le contrôle central est quelque chose qui réunit les espèces, le cerveau, au travers de son système
hypotalamo-hypophysaire va être au cœur de la régulation de la fécondation. Les productions
hormonales, GnRH (hypothalamus) et LH/FSH (hypophyse, sur ordre de GnRH), régulent le testicule.
La reproduction met classiquement en jeu 2 partenaires, un male et une femelle. Cependant
techniquement, on peut reproduire un seul individu.
Reproduction veut dire reproduire quelque chose. Chez les êtres inférieurs, on parle de
reproduction, la bactérie se reproduit à l’identique. On peut nuancer en disant reproduction non
sexuée. Mais elles peuvent aussi échanger des informations génétiques par la reproduction sexuée.
Donc se pose un problème dans l’emploi du terme reproduction chez nous. On n’est pas identique à
nos 2 parents, donc il faut se demander si ce terme est bon à employer chez nous. Evidemment non,
grâce à la méiose, on aura moitié du père, moitié de la mère, il y a aussi brassage génétique (crossing
over). Compte tenu du nombre de gène et du nombre d’échanges possibles, on considère que
statistiquement, on n’aura jamais 2 gamètes identiques. Chez la femme, c’est pareil, jamais un
ovocyte sera identique à l’autre. Le cas des jumeaux est très différent. Le meilleur terme à employer

Physiologie repro. et dev. animal Mr Richard Semestre 5
3
en ce qui nous concerne est « procréation » : création d’une nouvelle entité à partir de structures
existantes.
I. Physiologie féminines
A. Relation structure-fonction de l’ovaire
C’est un organe pair, plein, constitué
d’une région médullaire et d’une région
corticale. L’ovaire est, chez la femme,
systématiquement coiffé du pavillon de
l’oviducte, mais il n’y a pas de vrai contact
anatomique entre l’oviducte et l’ovaire. Du
coup on se demande comment l’ovaire
tient. On a un certain nombre de petits
ligaments qui vont l’accrocher dans la
cavité intra-péritonéale, et qui fait que
l’ovaire est bien maintenu en place. Cet
ovaire possède 2 fonctions, 1 fonction
exocrine, c’est la production de gamètes,
et une fonction endocrine, fabrication de
substances qui vont se retrouver dans la
circulation, ce sont les hormones, les plus
connues étant œstrogène et progestérone, mais il y a d’autres productions. L’ovaire dans son
fonctionnement, chez toutes les espèces, possède son cycle de fonctionnement. Chez tous les
organismes ce fonctionnement cyclique se réalise sur 2 périodes de la vie, à la puberté et jusqu’à un
arrêt de fonctionnement cyclique (différence entre humain et animal), la ménopause, qui est l’arrêt
de ce fonctionnement ovarien, et donc l’arrêt de la possibilité de procréation. Elle arrive à 35-40
ans, mais avec une très grosse variabilité.
La production de spermatozoïdes ne s’arrête normalement jamais. Potentiellement, il y a encore
production à 100 ans.
On trouve chez les vertébrés des animaux où l’on va trouver des organes creux. C’est le cas par
exemple chez les grenouilles. Le cas humain n’est pas le seul.
La région corticale, chez l’espèce humaine, est la région superficielle, à épaisseur variable, elle fait
de 2 à 10 mm, elle contient les follicules, et les dérivés folliculaires, que sont toutes les structures
après l’ovulation. C’est ce que l’on appelle corps jaune, corps blanc. Il reste toujours une trace, une
structure toute petite. L’épithélium de l’ovaire, constituant la périphérie, est en continuité avec
l’épithélium péritonéal, qui va former ce que l’on va appeler le ligament large, qui va s’accrocher au
péritoine, qui va être l’un des ligaments accrochant et reliant l’ovaire à la cavité péritonéale.

Physiologie repro. et dev. animal Mr Richard Semestre 5
4
La médula, partie centrale, est aussi
appelée hile, avec une bonne zone de
parenchyme, qui remplie cette structure,
et ce parenchyme va contenir les follicules
qui vont être ramenés à l’extérieur en fin
de vie, et on va aussi y trouver la
vascularisation. Dans chaque ovaire on
aura une grosse artère pénétrante, qui va
se scinder en petites artérioles qui vont
venir alimenter l’intégralité du tissu. On
aura un tissu veineux à l’autre bout
extrêmement présent. Cette
vascularisation va évoluer dans le temps, à
chaque cycle on aura modification de la
vascularisation. A chaque fois que l’on
aura le développement d’un follicule, on
aura développement cellulaire très
considérable, et donc il y aura nécessité
d’alimenter les cellules, et à chaque fois
que l’on aura un développement de ce
type, la vascularisation va suivre. Tout un
réseau vasculaire complet se met en
place. Il disparaitra quand le follicule va
régresser. Pour l’expliquer, il y a
fabrication par le follicule en croissance de
facteurs de croissance. Le plus
caractéristique est le VEGF (vascular
endothélium growth factor). Il induit la
fabrication de l’endothélium (capillaire). Le deuxième plus important est le TGF et ces systèmes de
croissance, produits par le follicule, attirent les capillaires, et autour de follicules le réseau vasculaire
se met en place. C’est typiquement un effet paracrine.
On a découvert cela en faisant des greffes d’ovaires, un peu partout, la greffe d’ovaire est très facile.
Cependant, il n’y a pas de développement ailleurs que dans l’utérus. La vascularisation se met en
place très rapidement, et la greffe prend aussi rapidement que la production de follicules est rapide.
Ces facteurs de croissance sont très largement aidés, en termes de production, par le couple LH/FSH.
L’activation génique se fait directement par la FSH.
On trouve aussi du tissu nerveux, il y a une vraie innervation de l’ovaire, on trouve en particulier des
nerfs du système orthosympathique. Dans l’ovaire, on trouve de la noradrénaline, de la dopamine,
du VIP, du NO. Le système est donc très particulier.

Physiologie repro. et dev. animal Mr Richard Semestre 5
5
1. Evolution du follicule
On trouve d’abord les follicules primordiaux qui donnent follicule primaire, secondaire, tertiaire,
etc. Tous les follicules sont présents dès le début de la vie de l’individu. Dès 6 mois de gestation, il y
a déjà tout de prêt, et déjà certains follicules dégénèrent. On considère qu’à la naissance il y a 250 à
500 000 follicules primordiaux par ovaire. Les ovocytes, dans cette situation, ont démarré leur
méiose, au stade fœtal, mais ils sont bloqués en prophase 1, c'est-à-dire au tout départ de la division
méiotique. Ce stock, à la naissance, ne grossira plus, et il va s’épuiser progressivement jusqu’à la
ménopause. On n’a jamais vu de rupture de stock avant la ménopause. Seuls 300 à 400 iront au
bout du développement. Le reste développe alors et va se nécroser, ou alors ils développent des
mécanismes d’apoptose, avant de démarrer le développement.
Le démarrage est le passage du follicule primordial au follicule primaire. D’une morphologie
aplatie, les cellules folliculaires prennent une forme cubique, pour organiser le follicule. Le follicule
primaire entraine une multiplication considérable des cellules folliculaires, qui vont s’assembler les
unes aux autres pour former une granulosa. A partir de là, l’ovocyte commence à croitre un petit
peu lui-même, prend le diamètre du follicule, qui atteint lui 180µm de diamètre. On atteint ensuite
le stade préantral, à 180µm, une petite couche cellulaire se forme autour de l’ovocyte, c’est la zone
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
1
/
54
100%