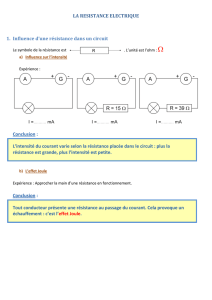
Notion de spéciation

Document préparé grâce "Au guide critique de l'évolution" sous la direction de guillaume LECOINTRE"
éd BELIN 2009
Doc. Spéciation
Une population ancestrale suit une expansion séparée par une zone inhospitalière (chaîne de
montagne, haut plateau, vallée désertique…) ; La population ancestrale se scinde de part et d’autre de
cette zone inhabitable. Les populations évoluent séparément sans qu’il existe réellement de frontière
nette, on observe plutôt un continuum de populations interfertiles et non une succession d’espèces
géographiquement bien délimitées. On peut au mieux distinguer des sous-espèces basées uniquement
sur des attributs comportementaux ou morphologiques.
Il n’y a qu’aux extrémités que coexistent deux espèces séparées génétiquement interstériles, sinon on
peut dire que partout ailleurs il n’existe qu’une seule espèce. Bien sûr au fur et à mesure de la
migration le flux de gènes est décroissant mais les populations restent interfertiles, même à travers
l’obstacle géographique (on le verra dans l’exemple du "faux anneau de spéciation des
salamandres californiennes ")
C’est en 1942, en étudiant les goélands de l’hémisphère nord que le biologiste E.MAYR a introduit la
notion de spéciation en anneau ; Il parla tout d’abord de recoupement circulaire puis en 1954, M.L
CAIN parle d’anneau de spéciation.
Le "faux anneau des salamandres" décrit par le généticien Georges.Ledyard STREBBINS
(1906-2000) cf.doc2
Un scénario datant de 1949 était classiquement admis:
Ensatina eschsholtzii oregonensis a entamé une expansion vers le sud il y a 7 à 10 Ma en
contournant la vallée aride de Joaquin en empruntant les deux versants boisés qui l'entoure.
Au cours de cette expansion, des différences génétiques apparaissent entre les deux branches
des anneaux (population côtière devient uniformément pigmentée et la population orientale
devient tachetée)
Six sous-espèces sont présentent à l'ouest; on observe un passage graduel de l'une à l'autre et
les hybridations sont possibles. En trois points des sous-espèces rentrent en contact en on
observe des hybridations ce qui prouve que l'éloignement génétique n'est pas important, on
observe seulement 8% d'hybridation, aucun cas d'hybridisme n'a été observé à ce jour; Les
deux sous-espèces divergent génétiquement et elles se comportent comme de vraies espèces.

Document préparé grâce "Au guide critique de l'évolution" sous la direction de guillaume LECOINTRE"
éd BELIN 2009
En 1998 de nouvelles données génétiques relatives aux salamandres ont été réanalysées et dès lors un
nouveau scénario apparaissait:
Certaines populations adjacentes présentent une divergence génétique très importante.
La succession des populations n'est pas toujours graduelle et certains hybrides présentent un
phénotype "forme et couleur" très différent.
Il est fort possible que les deux corridors boisés des versants de la vallée de Joaquin n'existaient
pas il y a 7 à 10 MA et donc que l'anneau géographique ne soit pas présent.
Au vu de ces faits, il est plutôt proposé qu'Ensatina eschsholtzii soit une super espèce,
regroupant environ 11 espèces de salamandres, qui aurait divergé par effet fondateur; C'est
seulement après que ces populations soient entrées en contact
1
/
2
100%