Acari, Ixodidea - INRA Montpellier

Acarologia is proudly non-prot,
with no page charges and free open access
Please help us maintain this system by
encouraging your institutes to subscribe to the print version of the journal
and by sending us your high quality research on the Acari.
Subscriptions: Year 2017 (Volume 57): 380 €
hp://www1.montpellier.inra.fr/CBGP/acarologia/subscribe.php
Previous volumes (2010-2015): 250 € / year (4 issues)
Acarologia, CBGP, CS 30016, 34988 MONTFERRIER-sur-LEZ Cedex, France
ACAROLOGIA
A quarterly journal of acarology, since 1959
Publishing on all aspects of the Acari
All information:
hp://www1.montpellier.inra.fr/CBGP/acarologia/
acarologia@supagro.inra.fr
Acarologia is under free license and distributed under the terms of the
Creative Commons-BY-NC-ND which permits unrestricted non-commercial use, distribution, and
reproduction in any medium, provided the original author and source are credited.
The digitalization of Acarologia papers prior to 2000 was supported by Agropolis Fondation under
the reference ID 1500-024 through the « Investissements d’avenir » programme
(Labex Agro: ANR-10-LABX-0001-01)

CONTRIBUTION
A
L'ÉTUDE
MORPHOLOGIQUE
ET
BIOLOGIQUE
DU
GENRE
APONOMMA
NEUMANN, r8gg (ACARIENS,
IXODIDEA)
I.
Ap.
flavomacttlatum (Lucas, r846).
II.
Ap.
latum (Koch, r844).
PAR
A.
SARATSIOTIS
*.
GÉNÉRALITÉS.
Les
Aponomma
sont
des espèces des régions tropicales
et
équatoriales d'Afrique, d'Asie
et
d'Amérique.
A
l'heure
actuelle le genre Apono1nma Neumann, r8gg est considéré comme très voisin
elu
genre
Amblyomma
Koch, r844,
par
la
communauté
de
la
plupart
des caractères morphologiques.
Cependant l'absence
d'études
approfondies
sur
toute
espèce d'Aponomma,
notamment
en ce qui
concerne
la
biologie, nous prive
d'arguments
complémentaires
pour
autoriser
cette
distinction.
C'est
NEUMANN
qui en r8gg 1 a séparé certaines espèces
elu
genre
Amblyomma
dans
un
nou-
veau
genre,
dont
les principaux caractères différentiels résident dans l'absence
d'yeux,
l'absence
de sillons
marginaux
(caractère valable à l'époque en fonction des Amblyomm.a connus)
et
l'as-
sociation normale avec les reptiles, à tous les
stades
. Une révision
systématique
de l'ensemble
du
genre
Aponomma
est d'ailleurs en cours de réalisation
par
le professeur
ANASTOS
et
ses col-
laborateurs.
Les
Aponomma
se scindent en
deux
groupes;
ceux à
scutum
coloré
par
des taches ou bandes
émaillées (groupe de
Ap.
exornatmn)
et
ceux a
scutum
non émaillé (groupe de
Ap.
latum). Les
espèces de
la
région australienne occupent une place à
part,
non seulement en ce qui concerne
la
coloration, mais aussi en ce qui touche certaines caractéristiques des sillons marginaux, épines
coxales, proportions, etc. ; ce groupe
pourrait
être
considéré comme une
entité
distincte.
I.
APONOMMA
FLAVOMACULATUM
Synonymies.
Jusqu'à
ces dernières années, A
p.
flavomaculatttm a été considéré comme une variation locale
et
extrême cl'Ap. exornatttm; en conséquence,
un
grand
nombre de références concernant
cette
*
Laboratoire
Vétérinaire
de
Thessaloniki-
66,
rue
de
26
Octobre-
Thessaloniki.
Directeur,
Mr
D.
YrAN-
NAKOULAS.
Travail
effectué
par
l'auteur
étant
expert
F.A
.O.
en
mission
officielle à
Pastoria-Kindia,
République
de
Guinée.
Acarnlo~ia,
t.
Xlii.
fasc. 3, 1972.

-477-
dernière
espèce
se
rapportent
en
fait
à A
p.
jlavomac1ûatum;
la
question
de
la
synonymie
exacte
des
références
anciennes
ne
peut
se
résoudre
que
par
le
réexamen
des
exemplaires
en
cause.
De
toutes
façons,
cette
synonymie
se
place
dans
le
cadre
des
dénominations
suivantes
Ixodes
flavomaculat~tm
Lucas, 1846.
Ophiodes
jlavomaculat~ts
(Lucas, 1846) Murray, 1879.
Aponomma gervaisi (Lucas, 1846) p.p. (Neumann, 1899).
Aponomma exornatttm (Koch, 1844)
p.p.
nombreux
auteurs).
Aponomma halli Tendeiro, 1849.
Aponomma
flavomac~tlatum
(Lucas, 1846) Tendeiro, 1955.
Distribution géogrophique.
Ne
sont
signalées
ici
que
les
références
concernant
avec
certitude
l'espèce
considérée.
Côte
d'Ivoire
:
MoREL
(1961) -Korhogo : Varanus
nilotic~ts,
Python
regi~ts;
Bouaké
: V. niloticus.
Dahomey:
MOREL
(1961) Agouagon : V. niloticus, V. exanthematicus, Python regius; Segboroué (Ouida) V. nilo-
ticus ;
Hountagbé
(Atiémé) : P. reghts.
Cameroun :
MoREL
& MoucHET (1958) Ndiki (Konn) : V.
nilotic~ts;
Ngaouyanga
: V. exanthematict,ts; Sakdgé :
V. exanthematicus.
MOREL
& MoucHET (1964) -Mekolo (Maroua) : V. nilotictts; Maroua : V. niloticus.
Centre-Afrique :
MoREL
&
FINELLE
(1961) -Ouaga (Bangui) : Python sebae; Soulemaka : V. nilotictts.
Guiné-Bissau :
TENDEIRO
(1948, 1950, 1951, 1955) -
Pessuba
: V. niloticus, V. exanthematicus ; Bissau : Python sebae,
Naja nigricollis;
Fulacunda
: V. exanthematictts;
Brene
(Bissau) ; V.
nilotiws;
Bissalanca (Bissau) :
V. exanthematictts;
Bambadinca
: V. nilotictts; Canchungo : V. exanthematic$tS; Pecixe :
V.
exanthe-
maticus;
Pessuba
: Python
sebae,
Naja nigricollis; Piche : V. exanthematicus;
Formosa
(Bijagos) :
V. niloticus.
Haute-Volta
:
MOREL
(1961) -Bobo-Dioulasso : V. exanthematicus, Python sebae;
Tiéra
(Bobo) V. exanthematic$tS;
Koriba
(Bobo) : V.
nilotic$tS
; Arli (Bobo) : V. nilotimts.
LAMONTELLERIE
(1962) -
Banfora;
Pô;
Ziniare;
Zorgo;
Garango.
Mali :
MOREL
(1961) -
Tombouctou
: V. exanthematicus;
Lorakbane
(Nioro) V. exanthematicus; Niono -
V.
nilotic$tS.
LAMONTELLERIE
(1960) -
Sanzana
: Python regius; Diou (Sikasso).
Mauritanie :
MoREL
(1961) -Rosso : V. exanthematicus; Boghé : V. exanthematicus.
Niger :
MoREL
(1961)-
Niamey:
V. exanthematic$tS;
Tilabéri:
V. exanthematicus;
Tlélahoua:
V. exanthematictts_
Sénégal :
LUCAS
(1846) ;
NEUMANN
(1899) -Sénégal : Varanus sp.
MoREL
(1961) -Malika (Dakar) : Crocodilus niloticus;
Dakar
: V. exanthematicus;
Hann
(Dakar) :
V. exanthematicus; Mbao
(Dakar);
Python
sebae,
V. exanthematicus;
Sangalkam
(Rufisque);
Naja

-478-
nigricollis, Python sebae; Bambey : Vara
mt
s
sp.;
Marsaloum (Thiès) : V. exanthematicits; Dara :
V. exanthematicus; Saint-Louis : V. niloticus; Podor : V. niloticus ;
Fatik
: V. exanthematicus;
Kolda :
V.
nilotimts; Niokolo-Koba : V. niloticus.
Sierra Leone :
MOREL
(rg6r) -Freetown.
Tchad:
MoREL
&
MAGIMEL
(1959)
-Fort-Lamy : V. niloticus V. exanthematici
tS
Pseudohaje goeldii.
Togo:
MOREL
(rg6r) -Sokodé.
Guinée :
MoREL
(rg6r) -Kérouané : Varanus sp. ; Ditine : V. niloticus ; Mamou : Python
sebae
; Kindia : Den-
droaspis viridi
s.
Références originales -Kindia : Dendroaspis veridis (4.IV.66;
I7.IV
.66; 30.IV.66;
rs
.v.
66;
27.V.66
;
IO.VI.66; I.VII.66; 22.VII.66; 5.
IX.66
; I6.rx.
66;
25
.IX.66; 8.X.
66;
20.X
.
66;
I.XI.66; 2I.XI.66 ;
rs.xn.66 ;
20
.I.67 ; 4.II.
67)
Dendroaspis jamesoni (4.IV.66; ro.vr.66; I3.
VII
.
66;
23.VII
.
66;
s.rx.66 ;
zs.rx.66 ;
rs.x.66;
r.xr.66; 23.XI.66;
rs.xn.66;
20.I.67; 4.II.67) Naja melanoleuca
(4.IV.66
;
rs.v
.
66
;
zr.vr.66;
r.vn.66;
r.rx.66; 5.x.
66
; r.xr.66; zr.xr.66; rs.xn.66) Python regius (3.m.66 ;
25.
rv.66; 7.vr.66; zz.vn.66; 8.x.
66)
Python
sebae
(7.vr.66) Naja nigricollis
(zr.m
.
66;
4.rv.66; zr.vr
66;
rs.vn.66;
22.VII.66; 4.Ix.66; I2.x.66) Varamts exanthematicus (s.vn.66). Il convient de faire
remarquer que l'abondance des récoltes sur serpents exprime l'intérêt qui s'attache aux captures
de
ces
reptiles à l'Institut Pasteur de Kindia, tandis que les hôtes normaux
de
l'espèce,
les
varans,
n'offrent pas d'intérêt médical.
Morphologie d'Aponomma flavomactûatum.
L'étude
morphologique a
été
effectuée
sur
la
descendance
d'une
femelle gorgée récoltée à
Kindia
sur
NaJa nigricollis
(zr.m.66),
de
même
que les observations biologiques.
Mâle (PL A).
Tiques de
petite
taille, de couleur
variant
du
brun-rouge
au
brun-noir. Les dimensions
moyennes
sont
les suivantes :
longueur
et
largeur
du
conscutum : 2,450
mm
X 2,630
mm;
longueur
du
capitulum
ventral
(base + hypostome) : r mm.
Basis capituli plus large que longue (ne
dépassant
pas
une fois
et
demi
la
longueur) ; bord
postérieur de
la
basis à cornes basidorsales bien formées, en angle
obtus
; bords
latéraux
de
la
basis réguliers, arrondis, sans saillies, de
contour
approximativement
rectangulaire. Palpes longs,
filiformes, à 4 articles
bien
individualisés ; en
vue
dorsale, article I
dépourvu
de soies, article
II
à 3 soies internes
et
3 externes, article
III
à 5 soies internes
et
2 externes ; en
vue
ventrale, article I
à 2 longues soies
internes
et
r
petite
externe
, article
II
à 4 soies internes
et
r externe, article
III
à 2 soies internes
et
r externe.
Face
ventrale
de
la
basis
nettement
rectangulaire, à 2 cornes basi-
ventrales
saillantes en angle aigu ; à
la
base de
chaque
corne, 3 épines de chaque côté. Chélicères
bien développés, leurs gaînes
portant
une
forte denticulation
jusqu'à
la
base,
diminuant
progres-
sivement
d'avant
en arrière.
Hypostome
à
dentition
bien marquée, à
3/3
files longitudinales
sur
8 rangs
transversaux
; coronule bien développée ;
partie
basale de
l'hypostome
longue.
Face
dorsale. Scapulae
très
saillantes,
en
angle
droit
ou
obtus
; sillons cervicaux courts
et
profonds, en virgule ; sillons
marginaux
absents
; festons uniformes, larges
et
courts, à soies sen-
sorielles. Yeux absents.
Sur
le conscutum,
la
distribution
des zones émaillées est
en
général celle
représentée
sur
la
figurer
(pl. A), avec des
variations
locales individuelles ;
ponctuations
de 2
types
;

-479-
ponctuations
grandes, disposées régulièrement,
non
sétifères ;
ponctuations
grandes sétifères,
sensorielles ;
un
peu
en arrière
du
centre
du
conscutum, 2 fossettes sensorielles à 7-8 épines, les
foveae centrales.
Face
ventrale. Gonopore mâle
au
niveau
des coxae
II,
à 2 lèvres tangentes. Uropore à valves
présentant
5 paires de soies
dont
3 postérieures
et
2 latérales.
Stigmates
en virgules, à partie cau-
dale régulièrement amincie, à courbure de
grand
rayon;
cadre chitineux régulièrement épaissi
2
mm
0,5
mm
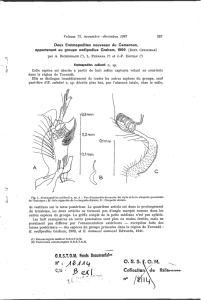
PLANCHE
A :
Ap.
flavomaculatum
c!.
I.
-
Conscutum
;
2.
-
Capitulum,
face
dorsale;
3· -
Capitulum,
face
ventrale;
4·
-
Coxae
I-II-III-IV;
5· -
Tarses
I-II-III-IV
; 6. -
Stigmate.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
1
/
21
100%