La synthèse des protéines.

La synthèse des protéines.
Problème posé:
La fabrication des protéines se déroule dans le cytoplasme, grâce aux ribosomes.
L'information, elle, se trouve dans le noyau, sous forme d'un segment d'ADN.
L'ADN, c'est la notice de montage de toutes les protéines qui nous constituent.
Il doit donc exister un relais.
Lorsqu'une partie de l'information héréditaire présente dans le noyau doit être utilisée dans le cytoplasme en
vue de réaliser une protéine particulière, elle est transcrite en une copie temporaire selon un mécanisme
original. Localement, la double hélice s'ouvre, de manière à permettre la fabrication d'un brin
complémentaire au segment de l'ADN qui correspond au gène à traduire en protéine. Ce brin complémentaire
est bâtit à partir de nucléotides d'ARN, pour former une molécule d'ARN messager.
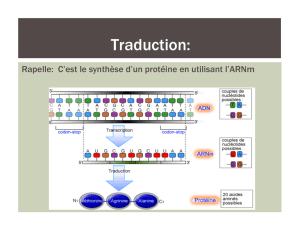
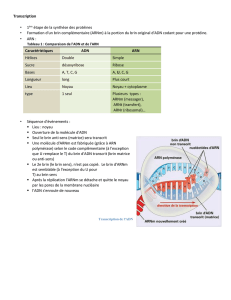
1 Transcription de l'information
Une protéine, l'ARN polymérase, est chargée de réaliser cette copie. Elle se glisse entre les deux montants de
l'échelle de l'ADN, qui s'écartent, séparant les paires de bases qui formaient les barreaux.
L'ARN polymérase rédige sa copie à partir de l'un des montants, elle lit les bases qui se succèdent et apparie
chacune avec une des bases complémentaires de nucléiotides d'ARN présents dans le nucléoplasme.
Ainsi une séquence d'ADN qui commence par AAATCGGCC... sera recopiée sous forme de
UUUAGCCGG... L'ARNm ressemble à une demi molécule d'ADN, une « échelle à un seul montant ».
Une fois la copie achevée, la molécule d'ARNm transcrite subit une maturation par:
- modification de ses extrémités (en vue de permettre l'attache au ribosome, d'un côté, d'éviter les
dégradations par les enzymes)
- épissage , élimination de certaines régions non codantes (introns) du gène transcrit
L'existence de ces introns chez les cellules eucaryotes surprend: quels peuvent être leurs rôles?
Les hypothèses les plus sérieuses avancent l'augmentation des recombinaisons de portions de gène (donc de
portions de futures protéines) lors de la méiose, en favorisant l'occurrence de crossing over, ou encore la
fabrication de différentes protéines à partir de la même séquence d'ADN transcrite, en modifiant les parties
conservées/éliminées de l'ARN brut.
La molécule ainsi formée, appelée l'ARN messager, quitte alors le noyau et passe dans le cytoplasme. Elle
amène « la notice de montage » d'une protéine, aux ribosomes chargés de sa fabrication.
2 Traduction de l'information
Une cellule connaît deux langages
celui que parle le noyau : le langage nucléique
celui que parle le cytoplasme la langage protéique
On passe de l'un à l'autre par une traduction.
Le langage nucléique Le langage protéique
s'exprime en nucléotides: 4 possibilités s'exprime en acides aminés: 20 possibilités
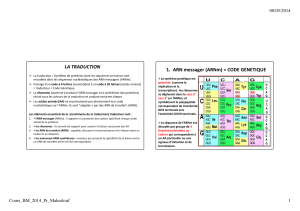
La correspondance n'est pas automatique: il faut imaginer qu'une combinaison de plusieurs nucléotides
détermine un acide aminé précis.
s'exprime par groupe de 3 nucléotides: le codon
Il y a 43 soit 64 codons possibles Il y aura trop de codons face au nombre d'acides
aminés, donc il existe des codons synonymes.
A chaque codon correspond un acide aminé particulier. Pour un même acide aminé il y a plusieurs codons
possibles. Le code génétique exprime la correspondance entre les codons et les acides aminés.
Compléments Synthèse des Protéines page 1 de 3

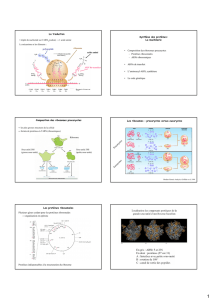
L'ARN messager s'encastre au sein du
ribosome.
Un ribosome lit les bases trois par trois (un
triplet d'ARNm est appelé codon).
Les ribosomes sont de très petits organites
(0,00002 mm) composés de 2 parties
globulaires. L'ARN messager s'encastre entre
les 2 globules du ribosome, puis défile sur toute
sa longueur, comme un fil de collier à travers
une perle. L'image est d'autant plus belle que
plusieurs ribosomes s'enfilent en même temps
sur un seul ARNm. Les ribosomes se
comportent comme des têtes de lecture: ils
orientent les codons d'ARNm afin de rendre
possible leur identification par un ARNt
complémentaire.
Les ARNt, t pour transfert, sont chargés
d'amener les acides aminés qui formeront la
protéine. Les ARNt possèdent d'un côté une
séquence de trois bases libres, et de l'autre un
acide aminé spécifique. Chaque molécule
d'ARNt porte à la fois un acide aminé et
l'anticodon qui lui correspond.
Les ARNt lisent le codon d'ARNm présenté par
le ribosome, et lorsque ce codon correspond à la
séquence de bases libres de l'un d'eux, tous
deux s'apparient temporairement, et l'acide
aminé qu'il portait est alors lié aux acides
aminés précédemment déposés...
Deux ARNt peuvent s'insérer en même temps
dans le ribosome; dès que le premier arrivé se
sépare de son acide aminé, en le liant à la suite
des autres, il quitte sa place et l'ensemble ARNt
et ribosome se décale le long de l'ARNm,
mettant à jour un nouveau codon, et la valse des
ARNt reprend jusqu'à ce qu'un codon STOP
signale la fin de la synthèse de la protéine.
Compléments Synthèse des Protéines page 2 de 3
2ème base
U C A G 3ème base
1ère base
Phénylalanine Sérine Tyrosine Cystéine U
UPhénylalanine Sérine Tyrosine Cystéine C
Leucine Sérine STOP STOP A
Leucine Sérine STOP Tryptophane G
Leucine Proline Histidine Arginine U
CLeucine Proline Histidine Arginine C
Leucine Proline Glutamine Arginine A
Leucine Proline Glutamine Arginine G
Isoleucine Thréonine Asparagine Sérine U
AIsoleucine Thréonine Asparagine Sérine C
Isoleucine Thréonine Lysine Arginine A
Méthionine Thréonine Lysine Arginine G
Valine Alanine Ac.Aspartique Glycine U
GValine Alanine Ac.Aspartique Glycine C
Valine Alanine Ac.Glutamique Glycine A
Valine Alanine Ac.Glutamique Glycine G

L'observation de cellules au microscope électronique a mis en évidence 2 populations de ribosomes:
ceux qui sont libres, et
ceux qui sont liés au réticulum endoplasmique rugueux.
Les ribosomes libres synthétisent surtout des protéines qui seront utiles au fonctionnement interne de la
cellule. Les ribosomes liés au réticulum endoplasmique rugueux synthétisent les protéines membranaires et
les protéines qui seront sécrétées à l'extérieur de la cellule.
Un même ribosome peut passer de l'état lié à l'état libre. La synthèse de toute protéine commencerait sur un
ribosome libre et ce serait la protéine en formation qui indiquerait au ribosome s'il doit se lier ou non au
réticulum endoplasmique rugueux. Les protéines sécrétoires sont marquées d'une séquence signal de 20 aa
environ qui permettent au ribosome de se lier au réticulum endoplasmique rugueux.
La fabrication d'une protéine requiert bien sûr de l'énergie ... fournie par les mitochondries.
3 Régulation de l'expression du matériel génétique (chez les procaryotes)
Le matériel chromosomique complet de l'organisme n'est pas exprimé totalement continuellement: il en
résulterait un gaspillage énergétique peu compatible avec la logique biologique.
On peut ainsi distinguer
•des gènes constitutifs, qui sont transcrits de façon permanente, comme les gènes codant les enzymes pour
le catabolisme du glucose
•des gènes de structure, qui sont transcrits selon des conditions particulières présentes dans
l'environnement à ce moment.
Citons aussi les gènes de régulation, dont qui fonctionnent selon deux logiques:
répression: contrôle - : une molécule inutilisée ou présente cesse d'être produite (gène de
l'anabolisme)
induction: contrôle + : une molécule utilisable déclenche la synthèse d'enzyme capable de la
dégrader (gène du catabolisme)
a) Répression
ex: le tryptophane, acide aminé, est synthétisé en 5 étapes à partir d'une molécule précurseur. Les
gènes des cinq enzymes qui dirigent cette synthèse sont situées côte à côte le long du chromosome
bactérien, avec le promoteur et l'opérateur. L'ensemble gènes de structure, promoteur et opérateur
est appelé opéron.
Ailleurs, il existe un gène régulateur qui est traduit en permanence et donne une protéine
« répresseur ».
Répresseur et tryptophane (=corépresseur) (si ce second est présent) peuvent se lier et s'accrocher
ensemble sur le gène opérateur, empêchant ainsi l'action de l'ARN-polymérase. Les gènes des
enzymes ne sont pas transcrits, et donc les enzymes, inutiles si le TRY est présent, ne seront pas
produites.
b) Induction
ex le lactose est un glucide qui peut être utilisé par la respiration cellulaire, moyennant une
transformation guidée par trois enzymes. L'opéron est constitué de la même façon, mais le
répresseur fonctionne différemment. Seul, il bloque l'opérateur, alors que si du lactose est présent
dans le milieu, le lactose se fixe sur la molécule de répresseur, et bloque son action négative sur la
transcription. Celle-ci peut se faire, les enzymes capables d'initier la dégradation du lactose sont
fabriquées, et le lactose pourra être utilisé.
Compléments Synthèse des Protéines page 3 de 3
1
/
3
100%