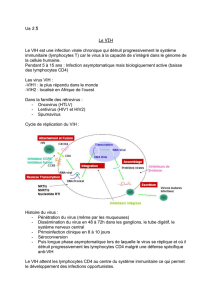
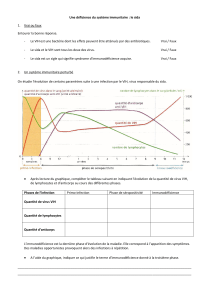
Étude du transfert du VIH-1 des cellules présentatrices d`antigènes

UNIVERSITÉ DE STRASBOURG
ÉCOLE DOCTORALE DES SCIENCES DE LA VIE ET DE LA SANTÉ
UMR_S1110 Interactions virus-hôte et maladies hépatiques
THÈSE présentée par :
Alizé PROUST
soutenue le : 20 septembre 2013
pour obtenir le grade de : Docteur de l’université de Strasbourg
Discipline/ Spécialité : Aspects Moléculaires et Cellulaires de la Biologie
Étude du transfert du VIH-1 des cellules
présentatrices d'antigènes aux
lymphocytes T CD4 primaires et
inhibition par les anticorps
neutralisants
THÈSE dirigée par :
Mme MOOG Christiane Directeur de Recherche, université de Strasbourg
RAPPORTEURS :
M. BRAND Denys PR-PU, université de Tours
Mme CHAKRABARTI Lisa Chef de laboratoire, Institut Pasteur de Paris
AUTRES MEMBRES DU JURY :
M. NEGRONI Matteo Directeur de recherche, université de Strasbourg

A mes Parents et
Aux trois autres Fantastiques
"To see a world in a grain of sand
And a heaven in a wild flower,
Hold infinity in the palm of your hand,
And eternity in an hour"
Auguries of Innocence, William Blake

REMERCIEMENTS

Remerciements
Comme le veut la tradition, je vais tenter de satisfaire au difficile exercice de la page des
remerciements. La difficulté tient dans le fait de n'oublier personne. C'est pourquoi je remercie par
avance ceux dont le nom n'apparaît pas ici et qui m'ont aidée d'une manière ou d'une autre.
Je tiens tout d'abord à exprimer mes remerciements aux membres du jury, Lisa Chakrabarti, Denys
Brand et Matteo Negroni pour avoir accepté la lourde tâche d'être rapporteurs de ce manuscrit et
ainsi d'évaluer mon travail de thèse malgré leurs emplois du temps chargés.
Un grand merci à Sidaction pour m'avoir financée durant ces trois ans. Sans votre association je
n'aurais pas pu réaliser ce projet.
Je remercie Catherine Florentz et Serge Potier qui ont répondu présents dans les moments les moins
faciles de cette thèse. Merci pour votre écoute et votre soutien. Les doctorants ont énormément de
chance de vous avoir.
Je remercie le Professeur Thomas Baumert de m'avoir accueillie dans un laboratoire d'excellence
comme il en n'existe peu en France.
Je remercie Christiane Moog pour m'avoir donné l'opportunité de travailler sur un sujet aussi
intéressant, important et actuel.
Je remercie également Vincent Holl sans qui j'aurais été perdue au début. Merci Vincent d'avoir
encadré mon travail durant mon stage de Master. Merci de ta confiance et de ton soutien durant
ces six premiers mois au laboratoire. Apprendre à travailler à tes côtés a été un réel plaisir. Merci
pour ton enthousiasme et ton dynamisme et pour toutes ces soirées où je t'ai fait partir à pas d'heure
car je ne comprenais rien à mes résultats de cytométrie.
Merci à Sylvie pour m'avoir tout appris sur les macrophages. Je suis bien loin de les connaître aussi
bien que toi mais je tiens à te remercier pour ta patience et ta confiance durant cette thèse. Merci
également pour m'avoir appris à "maniper" correctement.
Un remerciement spécial à Julien et Alex pour m'avoir appris (en plus d'autres choses bien plus
scientifiques) que j'étais un kinder: brune à l'extérieur, blonde à l'intérieur (comment aurais-je pu
savoir à l'époque que Milteniy était une marque et non un nom de famille ?!?).
Merci à Thomas pour ta bonne humeur constante, tes emails/sms alors qu'on est à 3 mètres l'un de
l'autre qui n'ont jamais cessé de me faire rire (ou au moins sourire) et ton écoute sans faille.
Je remercie Mélanie pour les lectures, relectures... re-re-lectures de ce manuscrit ou de toute autre
chose en rapport avec le laboratoire. Merci pour les déchiffrages des hiéroglyphes de la chef, pour les
excellents gâteaux (au moins tu sais que le salon de thé est envisageable pour la reconversion), pour
les ELISA et pour avoir été une voisine parfaite (bien que ma règle se soit souvent mystérieusement
translatée sur ton bureau !).

Remerciements
Merci à Bin et Marina pour les échanges de points de vue plus ou moins scientifiques. Pour leurs
accents (respectivement chinois et argentin) qui m'ont fait voyager sans bouger du labo. Marina, je
compte bien visiter un jour l'Argentine et t'y faire un coucou !
Merci à Gégé pour les fous rires. Je pense ne pas connaître d'autres personnes capables d'autant
parler de ses week-end que toi. Tu es la fraicheur incarnée.
Merci à Maryse qui elle aussi à subi les lectures et relectures de la thèse et de l'article.
Merci aux salles E309 et E308 (pièces "HCV") pour m'avoir accueillie lorsque l'évasion du bureau se
faisait nécessaire.
Je remercie les "jeunes" pour les bons moments de discussion à la cafet'. Mat, Catherine F, Lamine,
je vous souhaite bon courage. La fin est proche et je suis de tout cœur avec vous. Le bout du tunnel
sera plus long à atteindre pour Émilie et Sophie mais vous allez gérer ça comme des chefs. Les beer
times de fin de semaine se sont faits de moins en moins nombreux au fil des départs, à vous de les
relancer. Merci à Sarah, Laura et Charlotte pour leur bonne humeur de tous les jours. Sarah, ton
calme et ta patience m'ont souvent fait me demander pourquoi je m'énervais pour des choses
insignifiantes. Tu seras une maman merveilleuse pour "Isabello". Laura et Charlotte, votre caractère
"emporté" m'aura souvent fait rire. Vous vous complétez à merveille toutes les trois. Après les "4
Fantastiques" avons nous trouvé les "3 Mousquetaires" ?!?
Merci aux soutiens logistiques sans qui le labo ne pourrait pas tourner correctement. On vient
généralement peu vous voir mais vous faites énormément. Un grand merci à Sigis, Anne, Catherine
C, Jeremy, Patricia, Richard, Dominique et Halima.
En vrac mais loin d'être moins important: Merci au Roi des Sudoku Rajeev pour sa compréhension
et son inquiétude dès que j'avais un coup de mou. Merci au seul homme parmi toutes les femmes du
déjeuner : Éloi. Tu n'es pas là depuis bien longtemps mais ton intégration si rapide montre bien à
quel point tu es adorable et sympathique. Merci à Catherine S pour ses conseils avisés.
Un grand merci à mes amis hors labo (oui oui, il y a une vie en dehors du labo). Ceux qui ne savent
pas ce qu'est un P3 ou ce que signifie le terme "maniper". Greg, Mat, Karine, Vincent, Seb et autres
joueurs d'OdL, c'est une vraie bouffée d'air frais que de discuter avec vous. Évasion garantie vers
un "quelque part" bien loin de la thèse. Merci à Lionel pour les dépannages word/pdf, tu es un chef !
Un immense merci aux trois autres fantastiques: Laetitia, Isabel et Marine. Merci les filles pour
votre soutien inconditionnel dans les bons comme dans les moins bons (voir très mauvais) moments.
Merci pour votre bonne humeur sans laquelle ces trois ans auraient été plus que difficiles.
Marine, je ne pensais pas qu'un estomac pouvait supporter autant de chose. J'en reste encore
hallucinée.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
1
/
177
100%