DESTINEES DU PYRUVATE - medi

Destinées du pyruvate et cycle
tricarboxylique
M.E.H . CHERIFI
Cours de biochimie fondamentale 2014

Principales destinées du pyruvate
Dans les conditions
aérobiques le pyruvate
peut se transformer en
acide oxaloacétique;
un intermédiaire du
cycle de Krebs

Transformation du pyruvate en lactate
Réaction réversible anaérobique,se déroulant dans le cytoplasme
LDH est une enzyme ubiquitaire formée de 5 isoenzymes

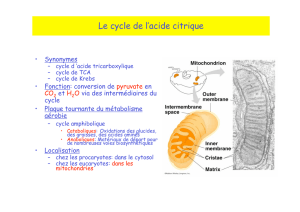
Passage du pyruvate dans les mitochondries
Cytoplasme
Matrice
Membrane interne
Mécanisme de symport de proton
Le NADH ne peut en aucun
cas traverser la membrane
mitochondriale interne.

Transformation du Pyruvate en Oxaloacétate
Cette voie n’est
pas la principale
voie suivie par le
pyruvate
intramitochondrial
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
1
/
42
100%