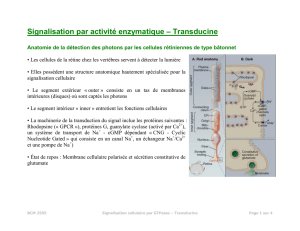
Les cellules en bâtonnet

LA PHOTOTRANSDUCTION
Certificat « organes des sens »
2005-2006
S. ALLOUCHE

RAPPELS
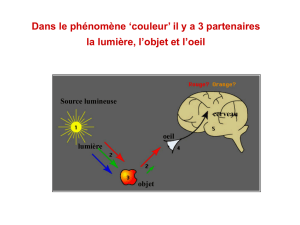
• L’œil reçoit la lumière (entre par pupille, est focalisée sur rétine par cristallin)
• Il est capable de l’absorber et de la transformer en énergie électrique (par l’intermédiaire
des cellules en cône et en bâtonnet de la rétine)
•Ce signal électrique est ensuite via le nerf optique transmis au cerveau qui reconstitue
l’image
• Ceci nécessite une coordination entre l’œil et le cerveau

1. Emission de rayons lumineux à partir d’un objet. Ces rayons sont focalisés sur la rétine et
forme une image inversée
2. Sur la rétine, les cellules photoréceptrices qui absorbent la lumière sont alors activées
3. Les cellules photoréceptrices envoient alors un signal électrique via le nerf optique jusqu’au
cerveau dans le cortex visuel
4. Ces signaux électriques reçus sont alors « décodés » et une image est alors reconstruite.
5. Il y a ensuite une interprétation de cette image reconstruite.

Comment se passe la transmission entre le signal lumineux et le signal électrique ?
• C’est au niveau des cellules photoréceptrices présentes dans la rétine que se fait cette
transformation de signal lumineux en signal électrique
La rétine
•Elle est constituée d'une fine membrane transparente qui tapisse toute la surface du fond de
l'œil et contient les cellules photo-réceptrices (sauf au niveau de la tâche aveugle)
•Sa structure : il y a donc plusieurs niveaux de cellules.
- une couche des cellules photo-réceptrices : les cellules en bâtonnet et les cellules en cône
(nommées ainsi de part leur forme).
- les neurones bipolaires constituent une nouvelle couche. En moyenne, 5 à 20 cellules
visuelles sont en relation avec un neurone bipolaire
- 5 à 10 neurones bipolaires sont en relation avec un neurone ganglionnaire (un million dans la
rétine) dont les axones constituent les fibres du nerf optique
• La lumière doit d’abord passer à travers les différentes couches de neurones avant
d’atteindre les cellules photo-réceptrices

La rétine
-l’épithélium pigmenté (mélanine) a
plusieurs rôles:
1. évite le reflet de la lumière vers les
cellules photo-réceptrices
2. Apport de vitamine A
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
1
/
54
100%
![B] Mécanismes de réception de la lumière par l`œil](http://s1.studylibfr.com/store/data/004008122_1-c8e3db34fa5cf688fb3a4102ae80c9b0-300x300.png)