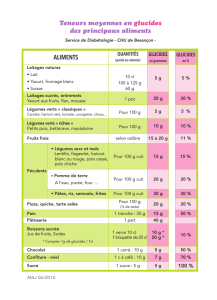
Les glucides dans la biologie des cellules.

Les glucides dans la biologie des cellules.
Les glucides sont des constituants organiques abondants chez les êtres vivants, et
notamment chez les végétaux. Ils comprennent par exemple la cellulose, qui est le composé
organique le plus représenté dans la biosphère, renfermant plus de la moitié du carbone
organique. Les glucides participent ainsi de manière essentielle à la réalisation des structures
du vivant. Mais, outre leur rôle structural, ils représentent aussi pour la cellule une source
d’énergie et des précurseurs pour de multiples biosynthèses.
Composés ternaires, ils sont représentés par un ensemble de petites molécules et de
macromolécules résultant de polymérisations.
Les glucides peuvent être importés dans la cellule, dans laquelle ils peuvent être
transformés en d’autres précurseurs glucidiques de petite taille ou polymérisés sous forme
de macromolécules, jouant des rôles divers.
Certaines cellules, dites autotrophes pour le carbone, peuvent réaliser la synthèse de
molécules glucidiques à partir du dioxyde de carbone. Cette synthèse nécessite de
l’énergie : celle-ci est la lumière dans le cas de la photosynthèse (cellule photolithotrophe),
l’énergie d’oxydation de certains composés minéraux dans le cas de la chimiosynthèse
(cellule chimiolithotrophe).
I / Les différents glucides et leurs rôles dans la biologie des cellules
On rappellera quels sont les principaux glucides intervenant dans la biologie des
cellules avec leur nature chimique et leurs propriétés en se limitant à celles qui sont
importantes dans le fonctionnement cellulaire.
1 / Les oses ou monosaccharides
- Formule générale : (CH20)n avec n compris entre 3 et 7.
- Aldoses et cétoses selon l’état d’oxydation du C1
- Formes linéaires et formes cycliques (cycle pyranne, furanne ; importance de la fonction
en C1, très réactive (cf. par exemple caractère réducteur, signification de la glycolyse).
- présence de carbone asymétrique (inutile de développer), si ce n’est ensuite pour
distinguer formes α et β dans certains polymères (amidon, cellulose).
- les plus fréquents : les hexoses : glucose, galactose, fructose, mannose ; les pentoses :
ribose et désoxyribose.
- On donnera les formules brutes et la représentation cyclique :
- du glucose (cycle pyranne) et du fructose (cycle furanne).
- du ribose et du désoxyribose.
- On donnera la formule développée du glycéraldéhyde.
- On mentionne l’existence et l’importance de dérivés glycosiques :
- acides aldoniques : oxydation de la fonction aldéhyde des aldoses.

- acides glycuroniques : dérivés carboxylés par oxydation du groupe hydroxyle
terminal.
- glycosamines : substitution d’un groupement amine primaire à l’hydroxyle en
position 2.
- On mentionne aussi la capacité à former des esters phosphorylés : grande importance dans
le métabolisme (ex : phosphoglycéraldéhyde).
- Intervention de nombreux oses dans les voies du métabolisme intermédiaire (cf. plus loin,
exemple : rôle important du phosphoglycéraldéhyde .
Précurseurs de polymères structuraux ou de réserves.
2/ Les disaccharides (ou diosides)
- La structure disaccharide résulte de l’union de deux oses par une liaison O-glycosidique :
schéma de la formation de la liaison.
- Principaux disaccharides, avec formules :
- maltose : liaison α (1->4) entre deux glucoses.
- lactose : liaison β (1->4) entre un glucose et un galactose.
- saccharose : liaison α (1->2) entre un glucose et un fructose ; schématisation de la
molécule ; modifications des propriétés découlant de la structure (caractère non réducteur,
relative inertie métabolique faisant du saccharose une forme de transport chez les végétaux,
forme de stockage chez certains végétaux).
3 / Les polysaccharides (ou polyosides)
- Polymères de monosaccharides sans taille moléculaire définie.
- Homopolysaccharides : ne contiennent qu’un seul type d’oses.
- Hétéropolysaccharides : contiennent différents oses ou dérivés glycosiques.
- Structures linéaires ou au contraire ramifiées.
- On présentera alors la structure et les propriétés de quelques polymères :
- glycogène : structure ramifiée : liaison α (1->4) avec tous les 8 à 12 résidus, une
ramification par liaison α (1->6). Cette structure fortement ramifiée permet une variation de
taille selon les besoins cellulaires. Présent dans les cellules animales mais aussi chez les
champignons.
- amidon : structures de l’α-amylose et de l’amylopectine.
- inuline : polymère de fructose chez les Astéracées.
On montrera en quoi ces polymères possèdent des propriétés en relation avec leurs
rôles de réserves : faible effet osmotique…(cf. plus loin).
- cellulose : liaison β (1->4), avec de nombreuses liaisons hydrogène assurant la
cohésion entre chaînes.

- chitine : enchaînement linéaire de N-acétyl-glucosamine. S’observe dans la paroi
cellulaire de champignons et de nombreuses algues. Grande importance dans la biomasse.
- les glycoaminoglycanes : enchaînements linéaires d’unités disaccharidiques de
taille moléculaire très variable. S’hydratent fortement ; nombreux groupements anioniques
qui font qu’ils ne se lient pas entre eux : les fluides riches en glycoaminoglycanes sont
visqueux. Constituants importants de la substance fondamentale du tissu conjonctif auquel
ils donnent une certaine turgescence. Adaptés à la lubrification des surfaces (articulations,
…) et à l’absorption des chocs (liquide synovial, cartilages…).
Exemple : acide hyaluronique (acide D-glucuronique et N-acétylglucosamine) ;
chondroïtine-sulfate (tissu cartilagineux) ; … héparine : grains de sécrétion de cellules
présentes à la surface vasculaire et abondantes dans les poumons, le foie, la peau. Inhibe la
coagulation du sang.
-les peptidoglycanes des parois bactériennes : enchaînements d’unités faites de N-
acétyl-glucosamine et d’acide N-acétylmuranique.
4 / Les hétérosides
La fonction hémi-acétalique d’un ose peut être condensée avec un groupe d’un
composé qui n’est pas de nature osidique. Formation de liaisons N-glycosidiques.
- Les nucléosides : hétéroglycosides de ribose ou de désoxyribose unis par une liaison β-
glycosidique avec l’azote 1 d’une pyrimidine ou l’azote 9 d’une purine.
- Les nucléotides : produits de phosphorylation d’un des groupements hydroxyles des
pentoses des nucléosides ; parfois structure cyclique par constitution d’une deuxième
liaison phosphoester avec un autre groupement OH du pentose (exemple : AMPc).
- On mentionnera la formation des dinucléotides, structures de base des coenzymes
d’oxydoréduction (NAD+, NADP+, FAD) et la polymérisation des nucléotides à l’origine
des acides nucléiques.
II / Origine des glucides cellulaires
1 / Production de glucides à partir de composés minéraux
a / la photosynthèse
Il s’agit de présenter la phase sombre de la photosynthèse avec le cycle de Calvin.
Approche expérimentale avec les expériences de marquage.
On indique comment la fixation du CO2 sur le ribulose 1,5 biphosphate conduit à la
synthèse de phosphoglycéraldéhyde : réduction endergonique du carbone du
phosphoglycérate grâce à l’ATP et au pouvoir réducteur NADPH.

On mentionne le devenir des trioses phosphates ainsi produits : régénération du
ribulose 1,5 biphosphate, synthèse d’hexoses, stockage temporaire sous forme d’amidon…
Les phénomènes seront localisés à l’échelle du chloroplaste puis replacés à l’échelle
de la cellule (sous forme de schémas).
b / la chimiosynthèse
Les enzymes du cycle de Calvin se trouvent dans certaines bactéries capables de
synthétiser leurs molécules organiques carbonées à partir de CO2. Chez les organismes
chimiolithotrophes, l’énergie provient de l’oxydation de composés minéraux.
On peut citer l’exemple de certaines bactéries s’inscrivant dans le cycle de l’azote,
comme celles de la nitrification (nitrosation, Nitrosomonas, et nitratation, Nitrobacter).
2 / Utilisation de glucides d’origine extracellulaire et entrée dans les cellules
a / Les sources des molécules glucidiques
Dans les cellules hétérotrophes, les glucides parviennent aux cellules sous forme de
petites molécules (nutriments). Celles-ci peuvent résulter de phénomènes de digestion des
glucides alimentaires (animaux, champignons, bactéries avec l’existence parfois
d’exoenzymes).
Au sein du végétal chlorophyllien, les cellules hétérotrophes des organes puits
utilisent notamment le saccharose, sécrété dans la sève élaborée au niveau des organes
sources.
Les molécules glucidiques peuvent aussi provenir d’une mobilisation des glucides
de réserves (cf plus loin).
b / L’entrée dans les cellules
Il convient ici d’envisager les structures et les processus qui assurent l’entrée des
glucides dans les cellules.
On pourra se fonder sur quelques exemples :
- étude de l’absorption du glucose dans la cellule intestinale : mise en évidence des
processus impliqués : entrée par transport actif secondaire, couplé au gradient de sodium ;
sortie par diffusion facilitée.
- entrée du glucose dans la cellule animale (exemples : cellules musculaires et adipeuses) :
on envisage alors les transporteurs à glucose et leur mise en action (approche cinétique,
…) ; il importe de montrer que ces échanges membranaires de glucose peuvent être
contrôlés par des hormones (insuline), susceptibles de modifier le nombre de transporteurs
en surface, en agissant sur les voies de recrutement. On précise alors le cycle des
transporteurs.
- entrée du saccharose dans les cellules puits du végétal chlorophyllien : existence de

transporteurs.
III/ Les glucides et la mise en réserves
1 / Les formes de mise en réserve
On rappellera que le stockage est fréquemment réalisé sous forme de
macromolécules en relation avec différents caractères :
- un gain de place (les molécules isolées pouvant se repousser entre elles).
- un effet osmotique limité (puisqu’il dépend de nombre de particules).
- une inertie chimique du fait que les groupements réactifs sont fréquemment impliqués
dans les liaisons de polymérisation.
- une facilité de synthèse : une seule réaction itérative, ce qui limite le nombre d’enzymes ;
nécessité cependant d’intermédiaires activés.
- une facilité d’hydrolyse : au plan énergétique, pas d’apport nécessaire ; une seule réaction
itérative.
- non diffusibilité hors du compartiment.
La mise en réserve peut néanmoins s’effectuer sous forme de molécules de plus
petite taille : exemple : saccharose.
On pourra également mentionner la présence de mannanes au niveau de la paroi de
cellules de certains albumens, dits alors cornés (exemples : palmier-dattier : noyau de datte ;
ivoire végétal).
On décrira quelques structures cellulaires de stockage :
amidon : amyloplastes : origine et structure
glycogène : granules hyaloplasmiques ; associations de molécules de glycogène et
d’enzymes.
Saccharose et inuline : vacuoles.
2 / Constitution et mobilisation des réserves
On rappelle les mécanismes essentiels de la polymérisation : celle-ci nécessite un
apport énergétique sous forme de glucides activés.
Exemple :
- UDP-glucose pour la formation du glycogène
- ADP-glucose pour la formation de l’amidon.
Chaque monomère apporte l’énergie pour sa propre incorporation.
On rappelle que la constitution et la mobilisation des réserves sont contrôlées !:
exemple du fonctionnement de la cellule hépatique !: glycogénogenèse – glycogénolyse
(phosphorolyse).
IV / Les glucides, sources d’énergie et de pouvoir réducteur
 6
7
8
6
7
8
1
/
8
100%