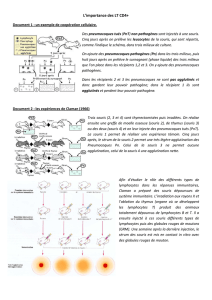
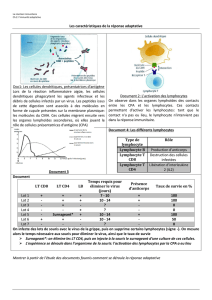
Rôle de la mutation vav1 r63w dans la sensibilité à l`asthme

Open Archive TOULOUSE Archive Ouverte (OATAO)
OATAO is an open access repository that collects the work of Toulouse researchers and
makes it freely available over the web where possible.
This is an author-deposited version published in : http://oatao.univ-toulouse.fr/
Eprints ID : 8642
To cite this version :
Bayol, Camille. Rôle de la mutation Vav1R63W dans la sensibilité
à l’asthme allergique. Thèse d'exercice, Médecine vétérinaire,
Ecole Nationale Vétérinaire de Toulouse - ENVT, 2012, 79 p.
Any correspondance concerning this service should be sent to the repository
administrator: staff-oata[email protected].

ANNEE 2012 THESE : 2012 – TOU 3 – 4080
RÔLE DE LA MUTATION VAV1R63W DANS LA
SENSIBILITE A L’ASTHME ALLERGIQUE
_________________
THESE
pour obtenir le grade de
DOCTEUR VETERINAIRE
DIPLOME D’ETAT
présentée et soutenue publiquement
devant l’Université Paul-Sabatier de Toulouse
par
BAYOL Camille
Née, le 10 Avril 1986 à RODEZ (12)
___________
Directeur de thèse : Mme Séverine BOULLIER
___________
JURY
PRESIDENT :
M Antoine BLANCHER
ASSESSEURS :
Mme Séverine BOULLIER
M Philippe GUERRE
MEMBRES INVITES :
M Hubert BRUGERE
Mme Anne DEJEAN
Professeur à l’Université Paul-Sabatier de TOULOUSE
Maître de Conférences à l’Ecole Nationale Vétérinaire de TOULOUSE
Professeur à l’Ecole Nationale Vétérinaire de TOULOUSE
Maître de Conférences à l’Ecole Nationale Vétérinaire de TOULOUSE
Docteur, Chargé de Recherche Première Classe à l’INSERM



 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
1
/
83
100%