Chapitre 5

Des colonies bactériennes lac+ et lac- sur un
milieu contenant un colorant indicateur
Chapitre 5
La génétique des bactéries et de leurs virus

Travailler avec des micro-organismes
Les procaryotes comprennent les
bactéries et les archéobactéries.
Les virus qui parasitent les
bactéries se nomment
bactériophages. À la différence
des eucaryotes, les organismes
procaryotes et les virus ont des
chromosomes simples qui ne
sont pas contenus dans l’espace
délimité par la membrane
nucléaire. Les procaryotes sont
haploïdes (ou monploïdes), ces
chromosomes ne subissent pas
de méiose, mais ils passent par
des stades qui rappellent ceux
de la méiose.

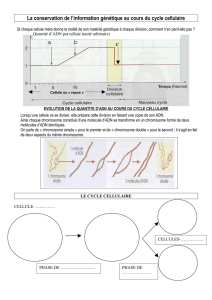
La réplication de l’ADN chromosomique bactérien
Les bactéries possèdent habituellement un seul
chromosome sous forme circulaire. Chez E. coli, la
réplication débute à partir d’une origine fixe (origine
unique appelée oriC) et procède ensuite de manière
bidirectionnelle (les fourches migrant vers les deux
extrémités du fragment en cours de réplication) pour se
terminer au niveau d’un site appelé site de terminaison.
Chez E. coli, un cycle de réplication-division ne dure qu’une vingtaine
de minutes.

Les types sauvages et les mutants
Les bactéries de type sauvage sont
prototrophes: elles peuvent former
des colonies sur un milieu minimum,
un substrat qui ne contient que
des sels inorganiques, une source
de carbone pour l’énergie et de l’eau.
Les clones (individus d’une colonie
descendant tous d’un ancêtre
génétique commun) mutants peuvent
être identifiés parce qu’ils sont
auxotrophes: ils ne poussent pas
tant que le milieu ne contient pas un
ou plusieurs nutriments spécifiques.
De plus, les organismes
de type sauvage sont sensibles à
certains inhibiteurs tels que la streptomycine, tandis que des mutants résistants
peuvent former des colonies malgré la présence de substances inhibitrices.

Les bactéries échangent de l’ADN par différents processus
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
1
/
28
100%