La vie dans la zone intertidale

Cours de Marie Gillon - 1
La vie dans la zone intertidale
La zone intertidale ou zone de balancement des marées, correspond à l’étage médio-littoral et
au supra-littoral celui-ci pouvant atteint par les marées de vives-eaux, y supportant une humectation
d’embruns rythmés par la marée.
Le milieu marin y subit les plus grandes variations de ses conditions physico-chimiques. Les
bandes côtières de l'Atlantique, de la Manche et de la mer du Nord passent alternativement du
domaine marin au domaine terrestre selon un rythme biquotidien auquel les espèces vivant dans cette
zone sont adaptées. La durée de l'exondation, liée à l'amplitude des marées, est le facteur majeur
agissant sur la vie dans ce milieu; elle y provoque dessication et déshydratation des organismes et en
détermine la température et la lumière. Les conditions de vie dépendent aussi de la nature du
substrat, différant sensiblement selon que celui-ci est meuble (graviers, sables, vases) ou dur
(roches).
I) Le cadre de vie
1) Un milieu dominé par la marée
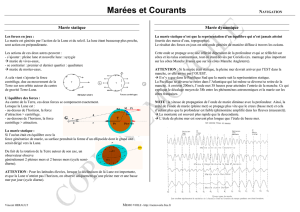
a) Fondements cosmiques et physiques de la marée
Le caractère cyclique de la marée résulte de la superposition de 3 composantes rythmiques,
chacune étant déterminée par des facteurs cosmiques (positions relatives de la terre, de la lune et du
soleil)
La première composante, dite " semi-diurne régulière ", est liée au mouvement apparent de la
lune et à l'attraction qu’elle exerce sur la pellicule océanique. C’est l'élément essentiel des
oscillations, les deux autres n'apportant que des modulations à ce mouvement principal. Elle se
manifeste deux fois par jour (période=12h25 ), l'arrivée de " l'onde de marée " en provenance de
l'Atlantique provoquant sur tout le littoral une élévation du niveau de l'eau (" flot " ou marée
montante) ; après une " étale de pleine mer" qui n'excède pas une heure, la mer se retire ("jusant" ou
marée descendante) pendant six heures et le cycle recommence.
D'une marée à l'autre, la variation du niveau de la mer (amplitude, ou marnage) est
sensiblement la même, mais le mouvement se décale progressivement de jour en jour car sa
périodicité est un peu différente de celle des jours solaires (24h50 au lieu de 24 h).
Le second rythme est sensible sur une durée de plusieurs jours et sa période coïncide avec celle
du cycle lunaire. Lors de la pleine lune ou de la nouvelle lune, l'amplitude des marées est la plus
grande (marées de "vive-eau"), elle passe par un minimum (marées de "morte-eau") au voisinage du
premier ou du dernier quartier. L'augmentation de l'amplitude constitue le " revif " de la marée, le
"déchet " (ou "baisse") correspond à sa diminution dont témoigne l'étagement des laisses de mer
successives, abandonnées chaque jour un peu plus bas sur le rivage.
Le troisième rythme est saisonnier. Lors des équinoxes, les marées atteignent des niveaux
exceptionnels; aux solstices, l'amplitude varie de façon plus discrète entre deux “vive-eau”
successives.

Cours de Marie Gillon - 2
Les variations de l'amplitude des marées se mesurent localement par la hauteur d'eau
subsistant au-dessus du niveau des plus basses mers possibles ("zéro des cartes"); le marnage de 4 à
5m en vive-eau moyenne sur les côtes atlantiques, variant d'un lieu à un autre en fonction des
conditions locales et surtout de la configuration du bassin
On a défini, pour quantifier les variations d'amplitude, une échelle théorique dont les valeurs
ou coefficients sont comprises entre 20 (amplitude la plus faible) et 120 (amplitude exceptionnelle).
b) Conséquences écologiques de la marée
Les oscillations périodiques du niveau de la mer délimitent sur l'estran une succession de
bandes parallèles au rivage, caractérisées par l'importance relative des durées d'émersion et
d'immersion.
Les organismes littoraux étant, pour la quasi-totalité d'origine marine, ils auront d'autant plus
de mal à survivre sur l'estran qu'ils seront émergés plus longtemps, à moins qu'ils ne disposent de
mécanismes perfectionnés leur permettant de résister pendant qu'ils sont hors de l'eau.
Dans les horizons supérieurs que la mer n'atteint que très exceptionnellement (coefficients
>115), le temps d'émersion est voisin de 100% ; seuls les embruns humectent le substrat, peu
d'organismes marins peuvent supporter les conditions y régnant. C'est la frange supra-littorale,
presque nue et pauvrement peuplée.
Au-dessous des pleines mers de vive-eau (coefficients voisins de 100, temps d 'émersion <
90%) commence l'étage médio-littoral: ses horizons supérieurs sont encore soumis à des exondations
importantes, mais l'ensemble de l'étage est richement peuplé ; ses habitants supportent bien
l'émersion mais ont besoin du retour régulier de la mer. Ils subissent quotidiennement de grandes
variations de leurs conditions de vie.
En bas de plage les émersions sont de plus en plus rares et de plus en plus brèves. Les
peuplements comportent des espèces mal protégées contre la dessication. C’est l’étage infra-litorral
dont seule la frange supérieure affleure pendant les basses mers de “vive-eau”. Il va en profondeur
jusqu’à 15-20m, tant que la lumière disponible est suffisante pour le développement d'un couvert
végétal dense.
c) Conséquences biologiques de la marée
L’étagement des peuplements littoraux: bien que le niveau bathymétrique des étages puisse
varier selon les paramètres locaux (agitation ou limpidité de l'eau etc.), ils confèrent aux rivages une
physionomie caractéristique.
La marée étant un phénomène rythmique, il en résulte des rythmes biologiques ou rythmes
tidaux liés et/ou associés à d’autre, correspondant à une alternance régulière d'activité et d'inactivité.
Celle-ci repose, selon les organismes, sur une simple réponse exogène au phénomène tidal ou sur un
rythme endogène.
On distingue alors plusieurs influences de la marée :
1. Rôle des vagues de pleine mer : Chez Littorina neritoides (Gastéropode à larve
planctonique et à vie adulte à l'horizon supralittoral), ce n'est que lorsque les vagues des pleines mers
de marée sizigiale arrivent à mouiller l'horizon supralittoral que les femelles déchargent dans l'eau
leurs capsules ovulaires.
Des populations de Littorines vivant à Plymouth, sur une jetée du port, en condition
d'immersion presque continue, suivent encore un rythme tidal, ne libérant leurs oeufs dans l'eau
qu'au moment des pleines mers, ce qui montre l'existence d'un rythme tidal endogène.
2. Rôle du rythme respiratoire en relation avec les marées : Chez de nombreux animaux
intertidaux, on a démontré l'existence d'un rythme respiratoire en relation avec les marées, la
consommation d'oxygène atteint son maximum en période de pleine mer. Ce rythme peut continuer
des semaines, voire des mois, lorsque les animaux sont placés en aquarium sans marée.
3. Rythme tidal simple : Le retrait des tentacules chez Actinia aequina (Tomate de mer)
semble du à une adaptation directe au rythme tidal. Le découvrement de l'animal par l'eau entraîne la
réaction protectrice de l'animal.

Cours de Marie Gillon - 3
4. Rythme tidal et rythme nycthéméral : Le rythme nycthéméral d'activité chez Actinia
aequina serait de type exogène : en effet, l’animal étend ses tentacules à l'obscurité, et devient inactif
à la lumière. Ce rythme serait un phénomène général sur lequel le rythme tidal s'inscrit en tant que
phénomène secondaire.
5. Coïncidence de rythmes tidaux sur la mobilité, la consommation d'oxygène, l'expansion
des mélanophores avec maximum pendant la basse mer : Chez plusieurs espèces américaines de
Crabes dont ceux du genre Uca, actives pendant la basse mer, on a démontré ces différentes
coïncidences. Tous ces phénomènes ont un maximum en période de basse mer, mais ils s'inscrivent à
leur tour sur des rythmes nycthéméraux, dont la coïncidence positive avec le rythme tidal peut
entraîner une exagération des phénomènes, tandis que leur rencontre en phase contraire peut les
estomper.
Chez les Uca dont les mélanophores s'étalent, leur couleur devenant plus foncée au moment de
la basse mer comme sous une luminosité intense, une basse marée diurne peut les conduire à avoir
une livrée foncée, une basse mer nocturne (ou une pleine mer diurne) détermine l'établissement de
teintes claires, mais non un éclaircissement maximal tel qu'il peut se réaliser lors de la pleine mer
nocturne. Ces interrelations traduisent l'écologie de ces Crabes à activité diurne et menant une vie
semi-terrestre.
D'autres Crabes intertidaux sont au contraire nocturnes et franchement aquatiques dans leur
écologie. L'activité de Carcinus moenas (crabe vert) se déroule surtout la nuit. Elle est exaltée si la
pleine mer survient la nuit, et déprimée tant en basse mer nocturne qu'en pleine mer diurne. Il faut
des semaines de maintien en aquarium sans marée pour que s'estompe un rythme d'activité non
seulement nycthéméral, mais dérivant chaque jour de 50 minutes environ, donc d'origine nettement
tidale, même pour des Crabes venant d'un bassin fermé pratiquement soustrait au jeu de la marée.
L'espèce méditerranéenne, Carcinus mediterraneus, vivant sur des côtes où la marée est peu
appréciable, ne présente au contraire que des rythmes nycthéméraux d'activité, à période de 24 h,
sans dérivation journalière de type tidal.
6. Rythme circatidal = rythme dont la périodicité est voisine de celle de la marées : Des
traitements thermiques permettent de réactiver un rythme endogène circatidal chez des Carcinus
moenas des côtes anglaises élevés en l'absence de marée. Le résultat est le même en replaçant ces
animaux dans la zone intertidale. Les mêmes expériences réalisées sur Carcinus mediterraneus
donnent des résultats différents: seul subsiste un rythme circadien. La spéciation a modifié à la fois
la morphologie et le fonctionnement de l'horloge interne.
7. Il existe souvent une interférences entre les deux types fondamentaux de rythmes
endogènes, circadiens et circatidaux chez des animaux et végétaux des zones de marées : Les
migrations verticales de la Diatomée psammicole Hanlzschia virgala se font verticalement. Elles
remontent vers la surface par basse mer diurne, mais jamais par basse mer nocturne. Ce
comportement serait dû à l'influence réciproque des deux rythmes.
La remontée des Diatomées (et d'autres microbenthontes) à chaque marée basse est
l'expression d'un rythme circatidal. Elle serait en partie inhibée pendant la nuit à cause du blocage
des migrations phototactiques dont la rythmicité est circadienne. Les manifestations phototactiques
positives sont à l'inverse exaltées le jour, mais la rythmicité circatidale a alors des effets inhibiteurs
qui s'opposent au développement de ces manifestations en dehors des phases de basse mer.
Le mécanisme synchroniseur du rythme est moins clair pour quelques Crustacés intertidaux :
c'est notamment le cas des Amphipodes et des Isopodes, habitant de fonds sableux. Ces espèces
parcourent la zone interdidale, en suivant la marée, c'est-à-dire qu'ils regagnent la plage avec la
marée montante pour s'enfouir dans le sable et recommencent à nager librement dès que la marée
descend, ce qui leur permet d'éviter de rester à sec dans la plage découverte. L’activité motrice de
ces Crustacés présente des maximums qui coïncident avec le début de la phase de marée basse.

Cours de Marie Gillon - 4
8. Rôle des mouvements des vagues : Chez l'Isopode Exocirolana chiltoni, particulièrement
sensible aux stimuli mécaniques, les mouvements des vagues exerceraient une action
synchronisatrice des rythmes d'activité.
Les Amphipodes ont une barysensibilité précise, et les variations de pression hydrostatique
joueraient le rôle de synchroniseurs. Chez l'Amphipode d'eau peu profonde Corophium volutator,
une diminution de la pression provoque l'activité de nage, plus actifs par basse mer, ils peuvent plus
facilement abandonner le substrat sur le point d'être découvert.
Chez d'autres Amphipodes, la réaction est inverse, c'est l'augmentation de pression
hydrostatique qui déclenche l'activité maximale. Cette réaction, dite “régulatrice” se retrouve chez
des benthontes d'eau peu profonde, placés dans des conditions éloignées de l'optimalité écologique
dans une eau trop profonde par pleine mer.
9. Influence synchronisatrice des variations de salinité : Chez des Crevettes Penoeidoe des
côtes atlantiques américaines, on a mis en évidence l'influence synchronisatrice des variations de
salinité. Le reflux est lié à une diminution de la salinité due à l'importance des eaux douces
continentales le long des côtes, et le phénomène inverse se produit à proximité de lagunes sursalées.
Chaque stade vital a sa propre halosensibilité, parfois capable d'apprécier des variations faibles
de l'ordre de 1/1000, ce qui permet aux premiers stades post-larvaires de gagner la zone côtière et
aux stades suivants de se déplacer vers le large.
10. Rythme tidal qui porte sur le taux de filtration de l'eau (exemple des Moules Mytilus
catiornianus et Mytilus edulis : non superposé à un rythme circadien, il est indépendant de la
température et la lumière. Au laboratoire, ce rythme persiste plusieurs semaines, en phase avec le
rythme de la localité d'où sont originaires les Moules. Transportés sur une côte à formule tidale
différente, leur rythme physiologique s'y adapte. Ce comportement semble la résultante de
l'adaptabilité d'un rythme endogène indépendant aux manifestations des rythmes des facteurs
environnants.
-Rm- Certaines exceptions apparentes peuvent s'expliquer sur des bases phylogéniques. Pour le
Décapode Callinectes sapidus ayant des rythmes tidaux semi-diurnes même lorsqu'il vit sur des
côtes où la marée suit un cycle diurne, l’explication se retrouverait dans le fait que la plus grande
partie de l'habitat typique de Callinecles s'étend sur des zones à marées semi-diurnes.
-chez Ostrea virginica, on aurait une influence synchronisatrice des phases de gravitation
lunaire et non de la marée elle-même. Si le rythme originel persiste longtemps quand les Huîtres
sont éloignées de leur grève, celui-ci montre ensuite une tendance à se synchroniser avec les phases
lunaires locales, même si les animaux sont loin de la côte.
2) Autres facteurs hydrologiques
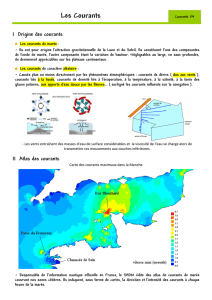
a) Les courants
Les masses d'eau déplacées au cours d'un cycle de marée sont considérables; elles engendrent
des courants littoraux alternatifs dont la puissance dépend du profil de l'estran et de la configuration
du rivage. Ils déterminent une importante dérive littorale et, dans les chenaux rétrécis (courant
d'Oléron, pertuis de Maubuisson, " passe d'Arcachon), ils sont si violents (2 m/s, soit 4 noeuds)
qu'ils déterminent une instabilité permanente des fonds sédimentaires. En revanche, ils s'étalent
paresseusement à l'abri des baies profondes.
La nature de la sédimentation côtière est déterminée par les conditions d'écoulement des
courants de marée car la mobilisation des sédiments dépend de leur énergie. Ainsi toute la côte
landaise est alimentée en sables bien calibrés provenant des alluvions de la Garonne qu'une
importante dérive littorale étale vers le sud.
En revanche, le ralentissement des courants au fur et à mesure qu'ils progressent dans le bassin
d'Arcachon s’accompagne du dépôt de sédiments de plus en plus fins vers la partie orientale.
L'affaiblissement des courants peut être provoqué par des obstacles discrets (promontoires littoraux,

Cours de Marie Gillon - 5
petites îles), par ramification des chenaux où remonte la marée, par la végétation du fond comme le
feuillage des Zostères). L'envasement plus ou moins important du substrat témoigne d'un
ralentissement sensible de l'écoulement des masses d'eau favorisant le dépôt.
b) Exposition aux vagues (= mode)
L'hydrodynamisme violent imprime aux peuplements des côtes exposées une caractéristique: il
est responsable d'un brassage important des eaux superficielles maintenant la température à des
niveaux assez élevés, favorisant la progression d'espèces thermophiles.
Les vents dominants poussent vers la côte des espèces pélagiques (Anatifes, Méduses,
Siphonophores, Vélelles...) s'échouant en grand nombre. Certaines, comme les Physalies,
proviennent d'eaux tropicales.
Toute réduction de l'hydrodynamisme favorise le réchauffement estival des eaux favorable à
l'ostréiculture activités ostréicoles. Le bassin d'Arcachon, la région d’Oléron sont des sites propices
au naissain.
3) Le substrat
Indépendamment de la nature lithologique ou de l'origine géologique des roches soulignant le
rivage, on distingue deux types de substrats en s'appuyant sur la nature des peuplements qui s'y
développent :
- les substrats " durs " susceptibles de supporter une flore algale et une faune fixée,
- les substrats " meubles " qui, en raison de leur structure et de leur mobilité, ne sont au
contraire pas capables de supporter une telle faune et flore.
Cette distinction s'appuie sur une évidence et souffre peu d'exceptions. Elle doit être nuancée
car les organismes vivants, en modifiant le milieu, conduisent parfois à des situations moins
manichéennes. Ainsi, la stabilisation d'un fond de graviers par les Zostères aboutit à la constitution
d'un biotope nouveau où se développe une végétation abondante attirant des Mollusques herbivores
(caractère de substrat dur), mais abritant une endofaune riche (caractère de substrat meuble).
L'estran ne présente pas une physionomie uniforme, les deux types de substrat peuvent
coexister sur une plage.
a) Les côtes rocheuses et leurs peuplements
Les conditions écologiques, liées aux marées, sont plus contraignantes pour les organismes des
zones à substrat dur. Sur les estrans rocheux, la zonation verticale de la flore et de la faune, très
marquée, dépend de la durée et de la régularité de l'exondation.
Les organismes, animaux ou végétaux, qui peuplent les estrans rocheux sont soumis à des
contraintes écologiques qu'aucune protection ne peut atténuer. Dans un 1e temps, ils supportent avec
rigueur les conséquences de l'alternance émersion-immersion. Cela entraîne donc des problèmes liés
à l'humectation, car l'eau ruisselle rapidement après le ressac.
On a de plus des problèmes liés à l'amplitude des variations thermiques pendant la marée basse
quand la température de l'eau de mer est remarquablement stable sur de longues durées. On a enfin
des problèmes liés aux variations de la salinité (pluie, évaporation...)
Les variations "en dents de scie" des conditions de vie se répercutent sur toutes les fonctions
physiologiques des animaux : respiration, économie hydrique, excrétion, possibilités de se nourrir...
La sévérité de ces contraintes, de plus en plus grande au fur et à mesure que l'on remonte l'estran, la
sélection des organismes en fonction de leurs possibilités d'adaptation aux conditions de la marée
basse, sera de plus en plus rude dans les horizons supérieurs.
En conséquence, la durée et la régularité de l’exondation conditionnent de façon très stricte la
répartition des organismes sur les estrans rocheux. Par exemple sur les côtes modérément exposées :
- la végétation souligne la variation continue des conditions d'humectation et se répartit en
horizons successifs, échelonnés de haut en bas de l’estran : lichens de la frange supra-littorale,
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
1
/
27
100%