D1-UE10-Diotel-Audition_et_systeme_vestibulaire_12.02.16

UE 10 : SYSTEME NEUROSENSORIEL
Nicolas DIOTEL
Date : 12/02/2015 Plage horaire : 8h30 – 10h30
Ronéistes : BOUREAU Laura Enseignant : Nicolas DIOTEL
ETHEVE Alixia
L’Audition et l’équilibre du système vestibulaire
I. Rappels et Introduction
1) L’audition et la fonction de l’audition
2) Les ondes sonores
3) Les caractéristiques du son
!
II. L’anatomie de l’oreille
1) L’oreille externe
2) l’oreille moyenne
3) l’oreille interne
!
III. Neurophysiologie auditive
1) Transmission du son aux cellules ciliées
2) Voies auditives
3) Modulation
!
IV. Neurophysiologie du système vestibulaire
1) Intro
2) Les organes à otholites : accélération linéaire
3) Les ampoules : accélération angulaire
4) Transmission des messages nerveux
5) Intégration
!
L’objectif de ce cours est de vous montrer que l’audition est un message sensoriel complexe (et que tout n’est pas
encore connu). Il faut connaitre le principe global de fonctionnement neurophysiologique de l’audition, et être
sensibilisé au fait que le bruit et les problèmes auditifs constituent un problème de santé publique majeur (ce
qui sera en fait plus détaillé dans le cours de sémiologie de l’audition). Il insiste en disant qu'il y a de plus en plus
de cas de surdité précoce lié à nos habitudes de vie (écouteurs trop forts, boite de nuit, concert). A partir d'une
certaine intensité sonore on aura un trouble de l'audition liée à une dégénérescence des cellules auditives, et
malheureusement on n'est pas encore capable de les régénérer.
!
!
! 1

I. Rappels et Introduction
!
Pour commencer, rappelons que le corps humain est régulièrement exposé à un grand nombre de stimuli.
Ces stimuli, bien que très différents en terme de nature et très variables en terme d’intensité sont pourtant
intégrés par le système nerveux central d’une façon très stéréotypée. En effet, les stimuli issus de notre
environnement seront détectés par un récepteur spécifique, cette activation entrainant la transduction du
signal. Cette transduction du signal est en pratique la transformation du signal sous sa forme initiale (par
exemple chimique ou physique) en un phénomène bioélectrique. Ce signal sera ensuite transporté sous
forme de potentiels d’action jusqu’au SNC. Ce SNC va venir intégrer ces informations de deux façons :
d’une part en y répondant directement de façon inconsciente via les réflexes (sensibilité inconsciente) ;
et d’autre part en traitant ces informations dans des centres spécifiques du signal. Ce ne sera que dans ce
cas qu’une sensation consciente de l’information sera perçue (sensibilité consciente) et qu’elle pourra
dans un second temps faire l’objet d’une intégration (en la comparant par exemple à des sensations
similaires déjà ressenties, faisant intervenir notamment les phénomènes de mémorisation, l’expérience
personnelle, et cætera…). On remarquera que c’est l’ensemble Sensation et Perception qui forment ce que
l’on appelle la Physiologie Sensorielle Subjective.
Cette physiologie sensorielle se fera toujours suivant 3 étapes :
(1) Réception et transmission de l’information au SNC – Conversion en message bioélectrique
(2) Analyse / intégration / interprétation de l’information
(3) Transmission d’une commande effectrice (ex: motrice ou autre).
!
! 2

On rappelle que l’entrée initiale de l’information
sensorielle se fait via des cellules spécialisées qui
réceptionnent les signaux: ce sont les récepteurs
sensoriels (neurone afférent). Ces récepteurs sont
connectés à des interneurones qui vont activer un
neurone efférent.
Ces récepteurs sont :
-soit liés à des neurones afférents
-soit eux-mêmes des neurones.
!
Dans le cas qui nous intéresse
aujourd’hui nous sommes dans le
premier cas de figure puisque les
cellules ciliées (qui détectent
l’information auditive) ne sont
pas des neurones et qu’elles sont
directement en relation avec le
SNC via des neurones. En
pratique, lorsque les stimuli
dépassent un certain seuil (on
rappelle qu’en dessous du seuil de
détection les cellules sont à leur
potentiel de repos), les cellules ciliées vont émettre un potentiel récepteur induisant ainsi la libération
d’un certain nombre de molécules chimiques au contact d’une terminaison d’un neurone permettant ainsi
la genèse d’un potentiel d’action.
Après ces rappels généraux concernant la physiologie sensorielle en général et les notions importantes de
potentiels d’action et de potentiels de récepteur, concentrons-nous sur l’objet de notre cours d’aujourd’hui
: L’audition. Comme vous le savez, c’est via l’oreille que nous sommes capables de détecter des sons.
Mais cette capacité à détecter des sons n’est permise que grâce à l’anatomie particulière de notre oreille
interne. Une fois détecté par l’oreille interne, le message auditif (qui est maintenant un message nerveux)
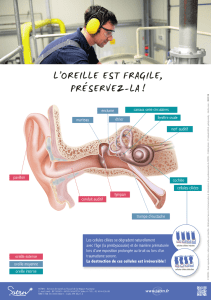
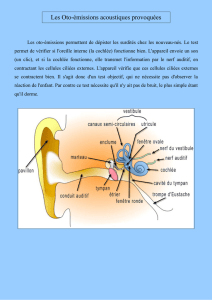
transite via le nerf Auditif jusque dans le SNC où il sera interprété. On remarquera que l’oreille interne est
constituée de deux parties : d’une part de la Cochlée et d’autre part de l’Appareil Vestibulaire (lui-même
constitué des Canaux semi-circulaires, de l’Utricule et du Saccule) qui n’ont pas du tout la même
fonction.
! 3

!
1) L’audition et la fonction de l’audition
L’audition correspond à la perception de l’énergie du son. Le son correspond à la partie audible du
spectre des vibrations acoustiques. Mais il ne faut surtout pas oublier que le spectre au complet des
vibrations acoustiques est beaucoup plus large que la petite partie de ce spectre que nous sommes
capables de percevoir. Les ondes sonores sont perçues en tant que variations de la pression de l’air (à
savoir un enchainement de zones de hautes pressions et de basses pressions. On va le voir plus en détails
par la suite).
Ainsi tout ce qui est susceptible d’induire un mouvement de l’air sera audible. La réception se fait au
niveau des oreilles. Les nerfs auditifs vont atteindre le cerveau, transmettre l’info à l’aire dédiée et
générer la perception du son.
L’audition (qui va de pair avec la parole) sert :
- A communiquer essentiellement
- A se reproduire: chez le cerf et chez la mésange la femelle choisit le mâle qui fait le plus de bruit
- Alerter: le cri est différent pour alerter d’un danger chez les singes en fonction de l’animal qui les
menace (aigle, léopard, serpent).
- A défendre son territoire: les phoques passent des heures à «chanter» pour empêcher les autres males
de s’approcher de leur territoire.
- S’orienter et chasser: la chauve-souris, comme les baleines et les dauphins, grâce à l’écholocalisation
(fonctionnement de type sonar).
!
!
!
!
!
! 4

2)Les ondes sonores
Les ondes sonores correspondent à la progression de vibrations de l’air dues à l’alternance de zones de
Haute pression (les molécules de l’air sont comprimées, concentrées) et de Basse pression (les
molécules de l’air sont raréfiées).
!
Le son correspond au final à une alternance de hautes et de basses pressions et tout cela va nous donner
une courbe sinusoïdale.
!
Le diapason est un excellent moyen de comprendre ce phénomène : quand il ne vibre pas les molécules
d’air qui l’entourent sont réparties de façon homogène. Dès qu’il vibre : il va comprimer d’un côté les
molécules d’air et au contraire provoquer la raréfaction des molécules d’air de l’autre côté, et ce de façon
séquentielle, provoquant l’émission d’ondes sonores successives dans les différentes directions de
l’espace (exactement de la même manière qu’une pierre jetée dans un lac qui génère des clapots
concentriques).
!
!
! 5
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
1
/
39
100%