Processus physiques et Stœchiométrie des transferts dans

0
Université de Rennes 1
Master 2 Ecologie Fonctionnelle Comportementale et Evolutive
Année 2009-2010
Mémoire Bibliographique
Processus physiques et Stœchiométrie des
transferts dans les écosystèmes
Pauline BRYERE
Sous la direction d’André-Jean FRANCEZ et Myriam BORMANS
Laboratoire : UMR 6553 Ecobio

1
Sommaire
Recherche Bibliographique : ................................................................................................... 2
Introduction : ............................................................................................................................ 2
I) Turbulence ............................................................................................................................ 4
1) ..................................................................................................................... Profondeur de mélange
.................................................................................................................................................... 4
2) Upwelling ............................................................................................................................... 6
II) Bioturbation ........................................................................................................................ 9
1)Effet de la bioturbation sur le ratio C : N de la litière ........................................................... 9
2) Minéralisation et infiltration de l’azote ............................................................................... 10
3) Impact de la bioturbation sur la disponibilité en phosphate ............................................... 11
4) Augmentation de la surface de l’interface eau-sédiments ................................................... 11
5) Effets des modifications de la disponibilité en nutriments par la bioturbation sur les autres
organismes ............................................................................................................................... 12
III) Changements globaux ..................................................................................................... 13
1)Effet de la hausse des températures en milieu terrestre ....................................................... 13
2)Effets conjugués de l’acidification des océans et de la température .................................... 14
3) Effets des UV sur la stœchiométrie ...................................................................................... 14
Conclusion……………………………………………………………………………………15

2
Recherche Bibliographique :
Mes recherches ont débuté par la définition de mots-clés nécessaires à la recherche
d’articles. Pour cela, je me suis aidée du diaporama fourni par mes maîtres de stage
concernant la stœchiométrie écologique et les relations trophiques. J’ai pu définir la liste
suivante : ecological stoichiometry, C : N : P ratios, light-nutrient hypothesis, nutrient
availability, bioturbation, upwelling. Cela m’a permis, par l’utilisation de moteurs de
recherches tels que Web of Knowledge et Google Scholar, de trouver une première partie de
mes références. Ces articles m’ont permis de trouver d’autres références. Mes maîtres de stage
m’ont également fourni un certain nombre de publications, notamment celles qui n’étaient pas
accessibles via les moteurs de recherches, ainsi que le livre « Ecological stoichiometry: the
biology of elements from molecules to the biosphere » (Sterner et Elser, 2002).
Introduction :
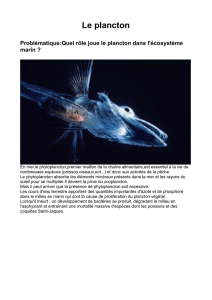
La stœchiométrie écologique permet une approche différente de l’approche classique
utilisée en écologie pour étudier les relations trophiques. En effet, alors que les interactions
entre les organismes et des organismes avec leur milieu sont souvent analysées à des échelles
allant de l’individu à l’écosystème, la stœchiométrie écologique propose une approche basée
sur les éléments chimiques. Cette discipline étudie ainsi l’équilibre des éléments chimiques
dans les interactions et les processus écologiques. Elle s’intéresse également aux flux
d’énergie et de matière au sein des écosystèmes (Sterner et Elser, 2002).
Les organismes vivants ont des compositions relativement constantes en divers
éléments tels que le carbone, l’azote et le phosphore. La plupart des études en écologie
stœchiométrique sont axées sur ces trois éléments et ce, depuis l’exemple de référence en
écologie stœchiométrique décrit par Redfield en 1958. Le rapport de Redfield prédit que le
ratio C : N : P du phytoplancton marin est constant pour toutes les espèces et est égal à
106 :16 : 1 (Redfield, 1958 in Sterner et Elser, 2002), une proportion égale à celle de la
photosynthèse. Ces proportions sont différentes et varient selon les milieux et les organismes
(Tableau 1). On peut observer une grande variabilité des autotrophes selon leur
environnement et leur besoins en éléments. Les plantes terrestres ont une composition plus
élevée en carbone, nécessaire à leur maintien et assurant une fonction de défense face au
broutage (Elser et al, 2000).

3
C: N: P
C: N
N: P
Références
Bruyère (toundra)
1943:33:1
59
34
Jonasson et al, 1999
Couvert forestier
1212:28:1
43.6±3.5
27.8±1.4
McGroddy et al, 2004
Sphaignes
tourbières tempérées
1986:32:1
59.9±17.0
33.5±10.8
Francez, non publié
tourbières boréales
2823:36:1
75.3±23
36.4±12.6
Pakarinen, 1977
Phytoplancton (eau douce)
166:20:1
10.0±3.0
30.0±16.0
Elser et al, 2000
Phytoplancton (milieu marin)
106:16:1
6.6
16
Redfield, 1958
Tableau 1 : Stœchiométrie des autotrophes dans différents écosystèmes.
Les rapports en éléments constitutifs des organismes sont différents le long de la
chaîne trophique : les herbivores n’ont pas les mêmes ratios que les plantes qu’ils mangent et
ainsi de suite en remontant dans les niveaux trophiques. McGroddy et al (2004) ont montré
que les rapports C : N : P (116 : 28 : 1) des herbivores en milieu forestier étaient nettement
inférieurs à ceux des végétaux. La différence est moins importante en milieu aquatique où ces
mêmes ratios sont plus faibles au sein des autotrophes. Selon les besoins des organismes,
certains éléments vont être davantage stockés ou recyclés. La régulation homéostatique
permet aux organismes de ne pas voir leur composition en éléments changer avec les
modifications du milieu extérieur. Kooijman (1995 in Sterner et Elser, 2002) définit
l’homéostasie comme « la capacité des organismes à garder constante la composition en
éléments chimiques de leur corps, malgré les changements de la composition chimique de leur
environnement, incluant leur nourriture ». Cependant, cette capacité diffère selon les types
d’organismes. Les autotrophes par exemple peuvent être plastiques concernant leur
stœchiométrie, principalement au niveau des rapports C : N : P (Droop, 1974 ; Ågren, 2004,
2008) et ce, en raison de leur capacité à stocker différents éléments en excès dans
l’environnement via une consommation de luxe (Klausmeier et al, 2004, 2008). Certains
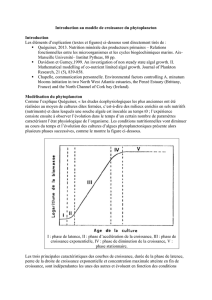
autotrophes sont d’ailleurs considérés comme non homéostatiques. C’est le cas notamment
d’une algue unicellulaire, Scenedesmus, dont le ratio N : P suit parfaitement celui des
ressources disponibles dans l’environnement (Figure 1 ; Rhee, 1978 in Sterner et Elser, 2002).
Les animaux en revanche montrent beaucoup moins de variations dans leur composition en
éléments et ont davantage tendance à être homéostatiques (Hall, 2009).

4
Figure 1 : Rapport N : P de l’algue unicellulaire Scenedesmus en fonction du rapport N : P du milieu. (A)
axes linéaire (B) axes logarithmiques. D’après Rhee, 1978.
La disponibilité en nutriments et leur stœchiométrie dans le milieu peuvent ainsi être à
l’origine de variations des rapports en nutriments (les rapports C : N, C : P et N : P étant
davantage développés dans les études réalisées) au sein des organismes, en particulier chez les
producteurs primaires. Différents facteurs peuvent être à l’origine de variations en éléments :
des processus chimiques (fertilisation, en agriculture notamment), biologiques
(décomposition, excrétion…) ou physiques. Il a en effet été montré que les différents ratios
des espèces végétales variaient en réponse aux différents apports de nutriments d’une part,
mais également aux apports d’énergie (Rastetter et al, 1997 ; Sterner et al, 1997). Tout facteur
susceptible d’avoir une influence les cycles biogéochimiques et sur la quantité de nutriments
ou de lumière disponibles aux organismes peut avoir un effet indirect sur leur stœchiométrie
ou leur croissance, notamment chez les autotrophes.
Ce rapport a pour but de montrer comment les processus physiques vont influencer la
stœchiométrie des ressources et des organismes et ainsi, par une redistribution des flux
d’éléments, le fonctionnement de l’écosystème. En particulier, nous examinerons les effets de
la turbulence et de la bioturbation. Les effets des changements globaux seront également
abordés.
I) Turbulence
1) Profondeur de mélange
La composition en éléments des autotrophes varie en réponse aux apports relatifs en
énergie et en nutriments majeurs (C, N, P) (Sterner et al, 1997). Tout processus ou facteur
affectant ces apports est susceptible d’entraîner des modifications dans la stœchiométrie des
producteurs primaires.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
1
/
23
100%