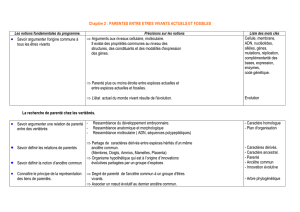
la lignée humaine : les homininés

PARENTE ENTRE LES ETRES VIVANTS ACTUELS ET FOSSILES
Les êtres vivants présentent une grande diversité mais aussi des unités de structure :
- la cellule
- l’ADN : support universel de l’information génétique
- similitudes entre les gènes de développement, ou gènes homéotiques
La composition des êtres est toujours la même : C – H – O – N. Il y a unité au niveau des plans
d’organisation, dans le fonctionnement : même molécule énergétique (l’ATP), même mode
d’expression des gènes, même code génétique, même mode de réplication semi conservatif de l’ADN.
Toute cette unité traduit une origine commune des espèces. Il y aurait un ancêtre commun à tous les
êtres vivants à partir duquel tous les êtres vivants actuels ou fossiles sont apparus après une série
d’étapes évolutives.
Tous les êtres vivants sont classés. L’unité de base est l’espèce (un groupe dont les individus sont
interféconds et partagent des caractères morphologiques).
LA RECHERCHE DE PARENTE CHEZ LES VERTEBRES : ETABLISSEMENT DE
PHYLOGENIE CHEZ LES VERTEBRES
On ne peut comparer que des caractères homologues (même organisation générale : membres en 3
parties, connexions identiques des structures avec le reste de l’organisme et dans le même ordre, même
origine embryologique…). Les processus évolutifs ont fait que des structures homologues peuvent
remplir des fonctions différentes (membre chiridien par exemple). Les espèces s’adaptent ainsi aux
différents modes de vie nécessaire pour occuper les diverses niches écologiques (fonction de l’espèce,
importance au sein d’un écosystème).
Certaines structures sont analogues mais pas homologues (différents types d’ailes n’ayant pas la même
origine embryologique).
I - Caractères macroscopiques cet parenté entre Vertébrés actuels
A. Comparaison de quelques caractères homologues
1) Caractères anatomiques ou morphologiques
a) Le membre chiridien
- construit sur le même plan chez tous les Vertébrés : 3 segments articulés : bras (un seul os, l’humérus
une seule pièce basale), avant-bras (2 pièces, radius – cubitus), main (nombreux carpiens,
métacarpiens, phalanges)
- membre attaché au reste du squelette par une ceinture : omoplate – clavicule
- tous les Vertébrés n’ont pas de membre chiridien : poisson
b) La structure de l’encéphale
- même structure de l’encéphale chez tous les Vertébrés : lobes olfactifs, hémisphères cérébraux, lobes
optiques, cervelet, bulbe rachidien alignés sur l’axe antéropostérieur
- différences sur la taille et la forme des segments
2) Caractères embryologiques
- ressemblance des stades embryonnaires des différents Vertébrés
- différenciation en fonction du milieu de vie
- mise en place très tôt du plan d’organisation similaire chez les Vertébrés
- ressemblances entre embryons parenté ?

- mise en place d’abord des caractères propres au groupe, puis au sous-groupe définition de taxons
de plus en plus restreints
3) Bilan
- ressemblances au niveau macroscopique hypothèse d’un ancêtre commun aux 5 classes de
Vertébrés (poissons, amphibiens, reptiles, oiseaux, mammifères ; une classe est un taxon)
- rapprochement entre les groupes grâce aux différences observées entre eux les différences et les
points communs entre les groupes renseignent sur la parenté entre les êtres vivants
B. Polarisation des caractères
1) Caractères primitifs / dérivés
- 2 états pour les caractères homologues : primitif (celui qui préexistait) et dérivé (celui qui est apparu)
- caractère dérivé : innovation évolutive (apparition d’un nouveau plan d’organisation, apparition,
disparition ou transformation d’un organe, apparition d’une nouvelle molécule)
- établissement de la phylogénie d’un groupe détermination des structures homologues héritées
d’un ancêtre commun aux différents taxons
/!\ Seul le partage de caractères dérivés témoigne d’une étroite parenté entre les taxons, on ne
peut pas établir de parenté grâce aux caractères primitifs.
2) Polarisation des caractères
a) Grâce aux données paléontologiques
- caractère dérivé = celui arrivé le plus récemment
- on peut choisir un extra groupe de sorte que l’ancêtre commun à l’extra groupe et au groupe étudié
soit plus ancien que l’ancêtre commun des espèces du groupe étudié si un caractère est présent chez
le groupe étudié et absent chez l’extra groupe, il est dérivé
- caractères de l’extra groupe : primitifs
b) Grâce aux données embryologiques
- au cours du développement embryonnaire, apparition d’abord des caractères primitifs puis dérivés
- partage de caractères dérivés liens de parenté étroits
C. Construction d’arbres phylogénétiques (cladogrammes)
- polarisation des caractères fabrication de matrices répertoriant les caractères dérivés et primitifs
des espèces étudiées construction d’un arbre phylogénétique (cladogramme) traduisant la parenté
entre les espèces
- construction de l’arbre :
∟ espèces d’un même âge au même niveau (à la même hauteur)
∟ nœuds : ancêtres communs (hypothétiques) aux espèces postérieures
- espèces les plus apparentés : celles qui ont un ancêtre commun exclusif non partagé par les autres
- groupes qui partagent un ancêtre commun exclusif : groupes frères
- ancêtre commun + tous ses descendants exclusifs : clade ou groupe monophylétique
∟ exemple : les Vertébrés (comprend toutes les espèces ayant un squelette interne, donc ayant
un ancêtre commun où le squelette est apparu)
∟ contre exemple : les Reptiles (Reptiles, tortue, lézard, crocodile ont un ancêtre commun
mais non exclusif qu’ils partagent avec les oiseaux les crocodiles sont plus proches des oiseaux que
des lézards)
- établissement d’un cladogramme fiable nécessité d’étudier un grand nombre de caractères
D. Espèces fossiles et ancêtres communs
- détermination des caractéristiques de l’ancêtre le plus récent grave au cladogramme : il a l’ensemble
des caractères dérivés partagés par les espèces qui lui sont postérieures
- mais pas de connaissance des autres caractères, donc impossible de l’identifier à un fossile trouvé
- impossible d’être sûr qu’un fossile est le premier être vivant à présenter une innovation évolutive, et
qu’il est l’ancêtre commun du clade

- établissement d’un cladogramme : on considère les fossiles comme des espèces actuelles
II - Caractères moléculaires et phylogénie
A. Notion de molécules homologues
- molécules appartenant à des espèces différentes ou présentes au sein d’une même espèce, semblables
dans leur structure mais présentant quelques variations, et n’ayant pas forcément la même fonction
∟ protéines homologues : variations dans la séquence d’acides aminés
∟ gènes homologues : variations dans la séquence de nucléotides
- établissement de liens de parenté entre les espèces possédant des molécules homologues en calculant
le % de différence entre les molécules
- molécules homologues différentes évolution des gènes qui les codent par mutation
- le nombre de mutations conservées est fonction du temps : nombre de différence ancienneté de
l’ancêtre commun possédant la molécule originelle
- liens de parenté par comparaison de molécules homologues chez des espèces différentes, mais
existence aussi de molécules homologues chez une même espèce (ex : globines chez l’Homme)
- molécules étudiées : celles qui ont une séquence une ressemblance de plus de 20% entre les
séquences ne peut pas être due au hasard
- pas de notion de caractères primitifs ou dérivés avec les molécules : on considère comme un
caractère la molécule entière, et non chaque acide aminé ou chaque base azotée
- la comparaison de molécules homologues ne permet pas de faire des clades
B. Des liens de parenté fondés sur des caractères moléculaires
1) Utilisation des molécules homologues
- on considère que :
∟ les molécules homologues dérivent d’une molécule possédée par le plus récent ancêtre
commun des espèces étudiées
∟ les différences entre les molécules homologues proviennent de mutations ; plus la molécule
ancestrale commune est ancienne, plus il y a eu de mutations
- si peu de différences entre 2 molécules homologues, les 2 espèces ont un ancêtre commun proche
- similitudes dans la séquence des molécules homologues similitudes dans les séquences
nucléotidiques des gènes homologues
∟ redondance du code génétique plus précis d’établir une phylogénie à partir des séquences
de gènes homologues qu’à partir des séquences de protéines homologues
2) Construction d’un phénogramme
- nombre de mutations à peu près constant dans le temps pour chaque protéine : apparition de
mutations périodiques, dont la fréquence dépend de la protéine ou du gène
- chaque nucléotide dans tout l’ADN a la même probabilité de muter
- taux de substitution matrice des distances avec % de différences entre les gènes établissement
de phénogrammes :
∟ comparaison des groupes 2 à 2
∟ longueur des branches proportionnelle au % de différence entre les groupes
∟ nœud : molécule ancestrale commune aux groupes situés aux extrémités des branches
∟ phénogrammes différents selon les molécules
/!\
- phénogramme degré global de ressemblance entre les molécules
- cladogramme liens de parenté
Ce sont des critères macroscopiques qui permettent d’établir des cladogrammes. Les cladogrammes ne
sont pas des arbres généalogiques puisque les ancêtres ne sont qu’hypothétiques. Ces cladogrammes
permettent de connaître les degrés de parenté et d’imaginer quelques grandes étapes de l’évolution des
êtres vivants. Il permettent d’établir une classification phylogénétique, reposant sur le partage de
caractères dérivés. Ces arbres se lisent en termes de groupes parents et non en termes d’ancêtre à

descendants, et cette classification évolutive regroupe les espèces en clades. Chaque cladogramme
peut évoluer avec la prise en compte de nouveaux caractères. Les caractères macroscopiques associés
aux données moléculaires permettent la classification phylogénétique des espèces actuelles.
Classiquement la taxinomie se fait sur le partage de caractères primitifs (les poissons ne sont pas un
clade, mais ils possèdent en commun des caractères primitifs : nageoires, écailles et vie aquatique).
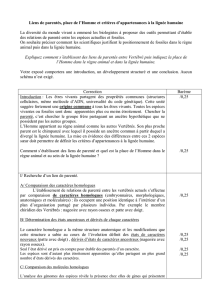
LA LIGNEE HUMAINE : LES HOMININES
I - La place de l’Homme dans le règne animal
A. L’Homme est un hominoïde
L’homme est un…
Caractéristique du clade
Date d’apparition du clade
Eucaryote
- 1.2 milliard d’années
Vertébré
Squelette interne
- 500 Ma
Tétrapode
Membres paires insérés sur
une seule pièce basale,
mâchoire
- 390 Ma
Amniote
- 340 Ma
Mammifère
Poils, glandes mammaires
- 220 Ma
Primate
Pouce opposable, ongle plat,
vision binoculaire
- 65 Ma
Hominoïde
Nez avec narines ouvertes,
vers le bas et rapprochées
- 23 Ma
Le clade hominoïde contient tous les grands singes anthropomorphes.
B. L’Homme est un hominidé
- gibbon : le seul hominoïde à avoir une musculature et un squelette de l’épaule totalement adapté à la
brachiation
- Homme – gorille – chimpanzé : groupe des hominidés
- données moléculaires, comportementales et chromosomiques très proche parenté entre les
Hommes et les panidés
- caryotypes chimpanzé et Homme : 99% du génome identique
C. Caractères de l’ancêtre commun à l’Homme et au chimpanzé
caractères dérivés homologues partagés par l’Homme et le chimpanzé
∟ bipédie occasionnelle
∟ utilisation d’outils rudimentaires après apprentissage
∟ vie en communauté
∟ arboricole
∟ 1m30, 40kg
∟ capacité crânienne : 350 – 400 cm3 (caractère primitif mais assez important)
- évolution des 2 groupes frères à partir de cet ancêtre en 2 lignées ayant des caractères propres à
chaque lignée
- divergence entre les 2 lignées : entre -10 et -5 Ma
- ancêtre hypothétique
II - Les critères d’appartenance à la lignée humaine
Toute espèce possédant au moins un caractère dérivé propre aux homininés fait partie
de la lignée humaine.

A. Caractères liés à la bipédie quasi permanente
- bipédie permanente adaptations anatomiques :
∟ colonne vertébrale à 4 courbures (cervicales – dorsales – lombaires – sacrées) centre
de gravité sur un axe vertical passant par la colonne vertébrale
∟ bassin court et aplatit donc large, en forme de cuvette, servant au maintient des viscères
dans le bassin ; bonne insertion des muscles grand fessiers pour le maintient du buste droit grâce à
l’élargissement
∟ col du fémur long, angle entre l’os et la vertical non nul pieds quasiment sous le
centre de gravité pendant la marche pas de dandinement
∟ genou épais amortissement des chocs
∟ trou occipital à la base du crâne, tête posée sur la colonne
∟ voûte plantaire développée, gros orteil dans l’alignement des autres (pas opposable), le
pied devient un organe de soutient et plus de préhension
∟ diminution du rapport des longueurs et du développement des membres antérieurs par
rapport aux membres supérieurs
- bipédie libération de la main pour l’activité locomotrice spécialisation dans la préhension
B. Caractères anatomiques liés au développement des capacités intellectuelles
- volume encéphalique : 1300 à 1450 cm3 (400 cm3 pour les autres hominidés)
- développement encéphalique développement du volume crânien
- boite crânienne ronde, développée vers le haut et l’arrière (« crâne cervical » ++, « crâne facial » --)
- face aplatie
- front haut ( développement du cortex frontal)
- plus de crête
- fosses nasales et menton alignés verticalement
- menton insertion des muscles labiaux ( parole)
- prognathisme disparu (plus d’avancée de l’os maxillaire) ← pas de développement du « crâne
facial », caractère juvénile conservé chez l’adulte, gène stoppé
- mâchoire en V
- dents de même taille, pas de crocs
C. Caractères liés à une activité sociale et culturelle
Tous les hominidés présentent une vie sociale en rapport avec leurs capacités intellectuelles, il existe
des rapports complexes entre les individus :
∟ capacité de domination
∟ capacité de jeu
∟ capacité d’apaisement
∟ capacité d’amitié
∟ capacité de trahison
∟ capacité d’alliance
∟ capacité de coalition
∟ capacité d’usage et de partage d’outils (même rudimentaires)
∟ capacité d’apprentissage aux jeunes par les adultes
∟ capacité d’imitation
Chez l’Homme, le développement de l’encéphale a permis :
∟ la progression des techniques, la fabrication d’outils sophistiqués et variés
∟ la maîtrise du feu
∟ le développement d’une pensée symbolique arts variés
∟ la manifestation de la conscience de soi et de la mort
∟ le sens du sacré, le rapport avec la religion, le divin, les offrandes et le culte des morts
∟ le langage articulé
Tout fossile présentant un de ces caractères dérivés anatomiques propres à l’Homme, associé ou
non à une trace culturelle, appartient aux homininés.
Hominisation = acquisition des caractères propres à la lignée humaine, de façon non linéaire
 6
7
8
6
7
8
1
/
8
100%