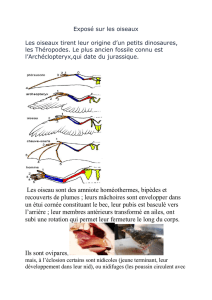
Introduction

1
Introduction
La Baie de Somme et la Plaine Maritime Picarde figurent parmi les espaces naturels français
les plus riches et les mieux connus sur le plan ornithologique. Dès 1833, une première mais
simple liste des espèces pouvant être rencontrées dans l'arrondissement d'Abbeville, unité
administrative englobant la Plaine Maritime et l'arrière-pays, était publiée (BAILLON, 1833).
Ultérieurement, un ouvrage plus ambitieux portant sur la même aire géographique apporte
des renseignements précieux sur l’avifaune locale au XIXème Siècle (MARCOTTE, 1860).
Ces travaux furent suivis de différents catalogues d'espèces figurant dans des collections
d'oiseaux naturalisés. Très tôt, les chasseurs – ornithologues publièrent le résultat de leurs
chasses ou de leurs observations, ce qui permet de disposer d'aperçus sur les espèces qui ont
fréquenté cet ensemble géographique au cours des 150 dernières années. Ce n'est cependant
que depuis le début des années 1960 qu'un effort plus soutenu fut fourni. Le grand pas en
avant est sans conteste la création de la première revue d'ornithologie régionale, L'Avocette,
en 1977. Dès 1983, une première avifaune moderne de la Baie de Somme voyait le jour
(COMMECY & SUEUR, 1983a). La même année sortait une avifaune du Hâble d'Ault
(TRIPLET, 1983a). Ces deux travaux furent suivis de différentes études d'espèces et
d'espaces, renforçant ainsi notre connaissance de la richesse avifaunistique locale. En 1990,
une nouvelle version de l'Avifaune fut éditée (SUEUR & COMMECY, 1990a) qui fut sans
conteste la publication ornithologique locale la plus diffusée. L'année suivante sortait un
rapport sur la reproduction des oiseaux d'eau de la Plaine Maritime Picarde (MOURONVAL
& TRIPLET, 1991a) qui permit d'améliorer les connaissances sur cette phase importante du
cycle annuel des oiseaux.
Dans le même temps, des études plus intenses étaient menées simultanément sur l'Huîtrier-pie
et sur les Laridés, qui aboutirent l'une et l'autre à la soutenance de thèses de doctorat
(TRIPLET, 1989 ; SUEUR, 1993), campant définitivement l'ornithologie picarde dans la
modernité.
Celle-ci eut par ailleurs différents supports. Pas moins de quatre revues régionales ont permis
de publier les synthèses annuelles ainsi que les différentes observations nouvelles :
L'Avocette, Documents zoologiques, Picardie Ecologie et Avifaune picarde. Documents
zoologiques et Picardie Ecologie ont disparu. Seule Avifaune picarde, revue dans laquelle
différents auteurs des deux revues disparues se retrouvent, continue à publier annuellement
les observations réalisées en Baie de Somme, au Hâble d'Ault et globalement sur l'ensemble
du territoire picard.
A ces revues principales il faut ajouter le Bulletin de la Société Linnéenne Nord – Picardie
qui a permis de publier quelques synthèses et le Bulletin annuel de l’Association
Marquenterre Nature (sous des appellations quelque peu différentes selon les années) mettant
à la disposition des ornithologues les observations réalisés chaque année sur le site du Parc
Ornithologique.
L'Ornithologie de la Plaine Maritime Picarde se caractérise également par au moins deux
publications par an dans des revues nationales d'ornithologie ou d'écologie.
Il était donc tout à fait légitime de terminer le siècle avec une synthèse de l'ensemble des
travaux menés localement. L'avifaune qui en résulte aurait dû donc paraître en 2000. Assez
exceptionnellement, les moyens financiers nécessaires à la sortie étaient disponibles alors
même que les textes n'étaient pas prêts. En effet, dans le cadre du projet transManche "Baie
de Somme – Baie de Rye, deux baies pour un même environnement", il était programmé de
dresser un inventaire avifaunistique de la Baie de Somme, qui soit l'équivalent des différentes
publications traitant de ce sujet Outre-Manche. Cette avifaune était d'autant plus nécessaire
que le nombre de travaux publiés ici et là ne cesse de grandir, rendant difficile l'accès aux

2
connaissances pour de nombreux ornithologues souhaitant mieux connaître les oiseaux de la
Baie de Somme et les travaux menés sur ceux-ci.
Cette avifaune constitue donc une synthèse la plus exhaustive possible des connaissances
acquises sur les oiseaux de la Baie de Somme et de la Plaine Maritime Picarde. Certes,
comme dans les avifaunes de 1983 et 1990, figurent les données anciennes et les statuts
actuels. Mais ce travail comprend aussi une synthèse des études menées récemment sur
différents aspects de la biologie des espèces. Ceci a entraîné un déséquilibre entre les espèces
très bien connues et celles qui le sont beaucoup moins, mais ce déséquilibre doit servir de
base pour des études nouvelles. Tout n'est pas connu sur les oiseaux de la Plaine Maritime
Picarde. Des espèces et des sujets de travail sont encore à aborder et devraient offrir aux
ornithologues débutants des satisfactions autres que celles de simplement observer sans être
certain que le travail fourni ne se résumera qu'à un chiffre dans un tableau.
Car cette avifaune veut également prouver que des études approfondies peuvent être menées
sans grands moyens, avec un minimum de persévérance et d'application. Le compte à rebours
pour une synthèse encore plus complète, dans dix ans, commence donc avec la sortie de ce
travail.
Nous aimerions remercier nos collègues qui ont répondu présents à notre demande en sachant
à l'avance que les délais d'écriture très brefs que nous leur imposions les obligeraient à
d'importants sacrifices au cours de cette année 1999. Jean-Claude ROBERT, le premier
contacté, manifesta aussitôt son enthousiasme, malgré un emploi du temps professionnel des
plus chargés. Outre ses textes, Jean-Claude accepta de fouiller dans la littérature et de vérifier
données et dates d'un grand nombre d'espèces. Francis MONTEL a reçu la tâche ingrate de
devoir mettre en tableau toutes les observations de la plupart des espèces, tableaux sans
lesquels l'analyse des variations saisonnières et annuelles n'aurait pas été possible. Il a par
ailleurs assuré les premières relectures… et les dernières, avec des délais toujours plus courts.
Florent VIOLET, Pierre ROYER, Jacques BELLARD, Philippe CARRUETTE, Arnaud
SOURNIA, Cédric FAGOT, Michel URBAN et Anne BACQUEVILLE ont accepté le plan
de rédaction que nous leur avons imposé et les multiples modifications que nous avons
apportées à leurs textes. Isabelle HECQUET-GALINIER, Maryline GENCE, Cédric FAGOT,
Arnaud SOURNIA, Grégory CASSORET et Frédéric LEPILIEZ ont assuré l'intendance sans
laquelle ce genre de réalisation n'est pas possible dans d'aussi brefs délais. Alors que nous
rédigions, ils géraient les graphiques, la mise en forme, les relations avec les maisons
d'édition, le budget et bien d'autres tâches tout aussi passionnantes.
Nicole et Geneviève, Aude et Marine, Nathalie, Pauline, Hortense et Agathe, ont vu leur
compagnon et époux, leurs pères consacrer plus de temps à l'écran de l'ordinateur qu'à la vie
familiale au cours de cette année. Notre maîtresse commune nous a laissé bien peu de temps
et nous ne pouvons que leur dédier cette avifaune dont elles partagent une certaine forme de
réussite.
Ce travail n'aurait pas pu trouver sa concrétisation rapide sans la confiance qu'a accordée le
Syndicat Mixte pour l'Aménagement de la Côte Picarde à Patrick TRIPLET. A aucun
moment au cours de cette longue année de gestation, Jérôme BIGNON, président du
SMACOPI et Jean-Christian CORNETTE, directeur, n'ont remis en cause ce projet qui
pourtant a entraîné des modifications permanentes dans l'emploi du temps des agents chargés
de la gestion des milieux naturels au SMACOPI.

3
Dans le cadre du projet Interreg, de nombreux organismes ont apporté des compléments
financiers à l'aide européenne. Nous remercions Monsieur le Directeur Régional de
l'Environnement, Monsieur le Président du Conseil Régional de Picardie, Monsieur le
Président du Conseil Général, Monsieur le Délégué du Conservatoire de L'Espace Littoral et
des Rivages Lacustres, Monsieur le Directeur de l'Office National de la Chasse et Monsieur le
Directeur de GSM pour l'aide apportée à ce projet.
Au sein de ces Etablissements, nous voudrions remercier plus particulièrement Christine
BRUNEL, Claire BLIN, Elodie MARTELLIERE, Guillemette ROLLAND, Thierry
RIGAUX et Thierry HAUCHARD pour la confiance qu'ils ont manifestée à Patrick
TRIPLET lors du montage du projet Interreg.
Messieurs GILLARD et LESTIENNE autorisent la prospection des bassins de décantation de
Grand-Laviers, M. DELTOMBE la colonie d'Ardéidés de Boismont par François SUEUR.
Qu’ils trouvent ici tous nos remerciements.
Enfin, ce travail n'aurait pu voir le jour sans les milliers d'heures passées sur le terrain par des
dizaines d'observateurs dont la liste serait trop longue à établir ici. Nous voudrions en
particulier rendre hommage à Noël RANSON, fondateur du Groupe d’Etudes et de Protection
des Oiseaux en Picardie, véritable pépinière d’ornithologues attachés à la protection des
espèces et des espaces. Nos sorties sur le terrain, nos publications, notre compréhension de la
situation des différentes espèces ont été menées avec différents collègues qui, pour différentes
raisons, n'ont pas participé directement à la rédaction de cette synthèse, mais sont néanmoins
présents par nos publications communes. Isabelle VERIERE, Gilles BECQUET, Xavier
COMMECY, Hugues DUPUICH, Pascal ÉTIENNE, Catherine et Eric FOURCY, Jean-
Baptiste MOURONVAL, Jérôme MOUTON et Jean NOSAL ont partagé nos sorties et ou
leurs exploitations sous forme de notes, synthèses ou articles.
Patrick DUFOSSE et Didier FOURNIER, gardes respectifs de la Réserve Naturelle de la Baie
de Somme et de la Réserve d'avifaune du Hâble d'Ault, ont la tâche de préserver les richesses
que nous mettons en évidence. Sans eux, bien des espèces seraient en difficulté.
En dehors de la région, nos amis et collègues Michel Alexandre CZAJKOWSKI, Annie et
Guy DUHAMEL, Vincent SCHRICKE et Pierre YESOU nous ont toujours témoigné leur
soutien et ont contribué à certaines de nos études.
CHRIS, Dominique DELMOTTE et Arnaud le KERVERN ont fourni les dessins.
Puisse l’an 2000 apporter ce retour aux valeurs fondamentales de l’Ornithologie picarde
alliant sérieux des études, publication régulière des résultats et actions en faveur de
l’avifaune.
François SUEUR & Patrick TRIPLET

4
LA BAIE DE SOMME ET LA PLAINE MARITIME PICARDE
La Plaine Maritime Picarde correspond à un ensemble de terres basses, isolées naturellement
ou artificiellement de la mer par un cordon de galets et par des digues appelées localement
renclôtures. Elle s'étend au nord jusqu'à la vallée de l'Authie et au sud jusqu'à la vallée de la
Bresle. A l'est, elle est limitée par la base du plateau calcaire, tandis qu'à l'ouest la Manche
constitue une limite variable.
De par sa formation particulière, ensablement et conquête de milieux à grand renfort de
digues tout au long des siècles passés, et par les nombreuses rivières et cours d'eau qui la
traversent, la Plaine Maritime Picarde constitue une entité dans laquelle l'eau et la terre
semblent ne faire qu’un. La conjugaison du contexte géomorphologique et de l'action
ancienne de l'homme sur le littoral déterminent une variété considérable de milieux naturels.
Se succèdent en effet, sur cet espace de près de 20 000 hectares, dunes blanches et grises,
prairies humides, marais tourbeux, bocage humide, plans d'eau... Une telle juxtaposition de
milieux variés est à l'origine d'une diversité biologique reconnue sur le plan international.
Au Nord, la Basse Vallée de l'Authie constitue une limite naturelle avec le département du
Pas-de-Calais. En amont, elle se présente comme une succession de marais au degré
d'atterrissement variable où domine la roselière colonisée par une strate arbustive. Subsistent
encore plusieurs prairies méso-hygrophiles dont certaines sont plantées de peupliers.
L'essentiel de cette zone est dévolu aux activités cynégétique et pastorale.
La Baie d’Authie couvre environ 2 000 ha et présente de nombreux points communs avec la
Baie de Somme, sur le plan de la richesse écologique, tout comme hélas sur le plan de la
sensibilité à l'ensablement.
Au sud de l'embouchure de l'Authie, les renclôtures (polders locaux) plus ou moins humides
s'étendent sur 500 hectares et se partagent entre pastoralisme et chasse, cette dernière activité
étant à l'origine des nombreuses mares de ce secteur.
A l'Ouest, le Marquenterre est séparé du milieu marin pour moitié par le massif dunaire dans
lequel subsistent quelques pannes humides et par une longue digue de protection. Le centre
de cette micro-région (un des "pays" picards) parcourue de canaux et de fossés et parsemée de
nombreuses mares de huttes est largement dévolu à la céréaliculture et offre donc un paysage
très ouvert. Plusieurs marais tourbeux, pour la plupart ceinturés de prairies hydrophiles et
colonisés par une végétation semi-ligneuse et arbustive sont adossés à la falaise morte à l'est,
notamment au niveau de Rue. A l'ouest, les renclôtures témoignent de la colonisation
ancienne des terres sur l'estuaire. Parmi celles-ci, figure le Parc Ornithologique du
Marquenterre, site à vocation écologique et pédagogique d'une superficie de 200 hectares.
La Basse Vallée de la Somme, située au centre de la Plaine Maritime Picarde constitue un
autre ensemble original de renclôtures édifiées par l'homme dès le milieu du XVIIIème siècle.
Cet ensemble s'étend sur près de 1 400 hectares et l'influence lointaine de la mer y est révélée
par les nombreux chenaux anciens qui le parcourent. Le milieu apparaît comme une vaste
mosaïque de plans d'eau à franges marécageuses, dispersés dans un complexe de prairies
humides. L’activité cynégétique et l'élevage y prédominent largement sur les différentes
autres activités.
Le bocage, relictuel et localisé à Favières et Froise (cette dernière localité située sur les
communes de Rue et de Quend), est caractérisé par un maillage assez lâche essentiellement
constitué de saules têtards. Il délimite un patchwork de cultures intensives et de pâtures
hydromorphes. Une opération de remembrement est la raison de la diminution des haies

5
d'Aubépines qui ne manquaient pas d'intérêt pour de nombreuses espèces. De plus, la
graphiose a décimé les Ormes de plus de trois mètres de haut et cette espèce n'est plus
présente que sous forme d'arbrisseau. l'aspect bocager est déjà fortement déstructuré, ce qui
n'empêche nullement l'arrachage des dernières haies, tandis que peu d'initiatives locales de
replantation sont à souligner.
Pour ces zones arrière-littorales, les risques de drainage n'ont pas disparu totalement et des
sites importants peuvent encore subir un manque d'eau.
De leur côté, les marais sont de plus en plus colonisés par la végétation arbustive qui, faute
d'entretien, supplante des habitats rares comme les roselières.
Au Sud, les Bas-Champs de Cayeux forment une vaste étendue très plane résultant du dépôt
ancien de sables et d'argiles. Protégés des entrées marines par une digue de mer, ces milieux
naturellement humides sont en proie au développement d'une végétation arbustive et exploités
pour l'élevage bovin et dans une moindre mesure, pour la céréaliculture.
A l'extrême sud, les marais du Hâble d'Ault résultent de la fermeture d'une dépression. Au
centre de cet espace de prairies traversées de canaux, la réserve d'avifaune appartenant à
l'Office National de la Chasse, se présente comme un vaste plan d'eau ceinturé de roselières
au-delà desquelles s'étendent des pelouses graveleuses et des dépressions saumâtres. Ce site
contribue fortement à la richesse écologique des Bas-Champs et constitue un pôle d'attraction
pour les ornithologues et les amateurs de nature.
La Baie de Somme couvre 7 200 ha est correspond à l’emboîtement de deux estuaires : celui
de la Somme au Sud et celui de la Maye au Nord. La Baie de Somme s’ouvre sur la Manche
orientale entre la Pointe de Saint-Quentin au nord et celle du Hourdel, au sud, distantes de
5 km, et forme une échancrure de 12 km. Elle inclut une partie herbacée de 1 500 ha environ
et une zone sablo-vaseuse dont 3000 ha sont en réserve naturelle.
Sur le plan faunistique, autre qu'avifaunistique, il faut signaler la présence du Phoque veau-
marin (Phoca vitulina) pour lequel la baie de Somme est le site abritant la colonie française la
plus importante, ainsi que trois autres espèces relevant également de la directive Habitats : le
Triton crêté (Triturus cristatus), le Liparis de Loesel (Liparis loeselii) et l’Ache rampante
(Apium repens), toutes trois trouvées dans les dépressions humides ou inondées périphériques
de la Baie.
En Baie de Somme, un ensablement important conduit à une profonde modification des
peuplements d'invertébrés et, à terme, à un bouleversement écologique qui ne sera pas sans
conséquence sur les oiseaux et sur l'économie locale (diminution des possibilités de pêche,
moindre attrait sur le plan touristique). De nombreuses initiatives sont actuellement prises
afin de bien appréhender l'étendue des problèmes et, le cas échéant, proposer des mesures
compensatoires. Cependant, le mal s'accroît. La Spartine Spartina anglica, espèce végétale
pionnière et fixatrice du sable s'implante partout et obère lourdement sur l'avenir.
Dans le Nord de la France, les dunes présentent une grande importance sur le plan
géographique en occupant 91 des 170 km du littoral du Nord, du Pas-de-Calais et de la
Somme. Sur le littoral de ce département, elles s’étendent sur 2700 ha dont près de 2000 ha
d’un seul tenant, avec des hauteurs de 20 à 30 mètres pour les dunes les plus élevées. Toutes
les sous-unités dunaires sont ici représentées depuis la dune embryonnaire à Oyat
(Ammophila arenaria) à la dune arbustive dominée par l’Argousier (Hippophae rhamnoides).
Dans les dépressions, les mares d’eau douce permettent l’installation d’espèces végétales
hygrophiles et aquatiques.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
231
232
233
234
235
236
237
238
239
240
241
242
243
244
245
246
247
248
249
250
251
252
253
254
255
256
257
258
259
260
261
262
263
264
265
266
267
268
269
270
271
272
273
274
275
276
277
278
279
280
281
282
283
284
285
286
287
288
289
290
291
292
293
294
295
296
297
298
299
300
301
302
303
304
305
306
307
308
309
310
311
312
313
314
315
316
317
318
319
320
321
322
323
324
325
326
327
328
329
330
331
332
333
334
335
336
337
338
339
340
341
342
343
344
345
346
347
348
349
350
351
352
353
354
355
356
357
358
359
360
361
362
363
364
365
366
367
368
369
370
371
372
373
374
375
376
377
378
379
380
381
382
383
384
385
386
387
388
389
390
391
392
393
394
395
396
397
398
399
400
401
402
403
404
405
406
407
408
409
410
411
412
413
414
415
416
417
418
419
420
421
422
423
424
425
426
427
428
429
430
431
432
433
434
435
436
437
438
439
440
441
442
443
444
445
446
447
448
449
450
451
452
453
454
455
456
457
458
459
460
461
462
463
464
465
466
467
468
469
470
471
472
473
474
475
476
477
478
479
480
481
482
483
484
485
486
487
488
489
490
491
492
493
494
495
496
497
498
499
500
501
502
503
504
505
506
507
508
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
231
232
233
234
235
236
237
238
239
240
241
242
243
244
245
246
247
248
249
250
251
252
253
254
255
256
257
258
259
260
261
262
263
264
265
266
267
268
269
270
271
272
273
274
275
276
277
278
279
280
281
282
283
284
285
286
287
288
289
290
291
292
293
294
295
296
297
298
299
300
301
302
303
304
305
306
307
308
309
310
311
312
313
314
315
316
317
318
319
320
321
322
323
324
325
326
327
328
329
330
331
332
333
334
335
336
337
338
339
340
341
342
343
344
345
346
347
348
349
350
351
352
353
354
355
356
357
358
359
360
361
362
363
364
365
366
367
368
369
370
371
372
373
374
375
376
377
378
379
380
381
382
383
384
385
386
387
388
389
390
391
392
393
394
395
396
397
398
399
400
401
402
403
404
405
406
407
408
409
410
411
412
413
414
415
416
417
418
419
420
421
422
423
424
425
426
427
428
429
430
431
432
433
434
435
436
437
438
439
440
441
442
443
444
445
446
447
448
449
450
451
452
453
454
455
456
457
458
459
460
461
462
463
464
465
466
467
468
469
470
471
472
473
474
475
476
477
478
479
480
481
482
483
484
485
486
487
488
489
490
491
492
493
494
495
496
497
498
499
500
501
502
503
504
505
506
507
508
1
/
508
100%