le système nerveux sympathique

1
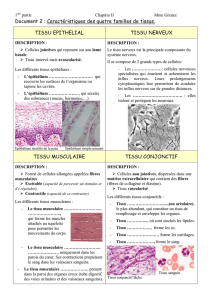
LE SYSTÈME NERVEUX SYMPATHIQUE
Historique
- Winslow (1716) : "Sympathia" en grec veut dire "souffrir avec" et lui donne pour rôle
de régir le monde intérieur (viscères).
- Langley l'appelle système nerveux autonome parce qu'il échappe à la volonté de
l'Homme.
- Muller (1840) parle du système nerveux cérébro-spinal pour les muscles striés et du
système nerveux autonome sympathique pour les muscles lisses.
Il est le plus ancien dans l'ordre phylogénique, il précède le système cérébro-spinal.
Définition
Le système nerveux, autonome ou végétatif assure l'homéostasie et l'équilibre du milieu intérieur :
motricité et sensibilité des viscéres, du coeur, du poumon, des vaisseaux.
régulation des sécrétions hormonales et glandulaires (système neuro-endocrinien).
sudation (glandes sudoripares) et piloérection des téguments.
Le système nerveux autonome organise la défense contre l'agression extérieure et assure la survie de
l'individu et de l'espèce. Il permet par des réactions viscérales adaptatives d'être le plus opérant lors
de l'agression. Il oriente le système nerveux cérébro-spinal vers une performance optimale.
Opposition du système nerveux cérébro-spinal et du système nerveux
sympathique
le système nerveux cérébro-spinal : gère les relations avec le monde extérieur.
Il intervient dans :
la sensibilité
extéroceptive
la motricité volontaire
la vie de relation (vie
psychique consciente).
Le système nerveux sympathique : régit lui le monde intérieur et tient sous sa dépendance la vie
végétative.
Il intervient dans :
l'équilibre intérieur et l'homéostasie
la motricité et la sensibilité viscérale
le système neuro-vasculaire
le système neuro-endocrinien avec les sécrétions hormonales et glandulaires.

2
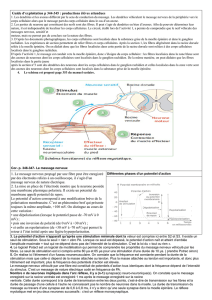
Opposition du système ortho et parasympathique
Le système nerveux végétatif est composé de deux systèmes que l'on a longtemps opposé mais qui
en fait ont une action complémentaire au niveau des viscères.
Le système orthosympathique
L'organisation est métamérique, horizontale et transversale :
Les centres médullaires sont étagés le long de la moëlle thoraco-abdominale.
Le neuromédiateur effecteur et la noradrénaline et sa synapse avec un ganglion modifié ( la
médullosurrénale) permet de libérer l'adrénaline. La voie humorale vient compléter la voie
nerveuse.
Il est le support de la sensibilité viscérale et des vaisseaux.
Les fibres pré-ganglionnaires courtes font relais dans une chaîne ganglionnaire latéro-
vertébrale proche de la moelle.
Les fibres post-ganglionnaires longues suivent les axes vasculaires et réalisent un large
maillage transversal étagé.
Ce système est le support des réactions de défense vis-à-vis de l'agression extérieure. Il est
dit ergotrope.
Le système para-sympathique
Les centres sont situés aux deux extrémités du névraxe (crânien et sacré).
L'organisation des fibres est longitudinale
Les fibres pré-ganglionnaires longues suivent le trajet des nerfs et font relais dans des
ganglions près des viscères (ganglions pré-viscéraux )
Le neuromédiateur est l'acétylcholine .
Le système est dit trophotrope , il gère les reserves énergétiques, est impliqué dans
l'exécution de fonctions (coeur,poumon,miction défécation &) . Il intervient dans la survie de
l'individu et de l'espèce.
Organisation générale
- Les centres axiaux sont médullaires pour l'orthosympathique, crâniens et sacrés pour le
parasympathique.
- Les afférences empruntent les voies de la sensibilité générale pour le système
orthosympathique les nerfs, pneumogastrique et glosso-pharyngien pour le parasympathique
crânien.
- Les efférences se composent d'un système à deux neurones, un neurone pré-ganglionnaire
ou système connecteur dont le médiateur est l'acétylcholine (récepteur nicotinique) et un
neurone post-ganglionnaire ou système effecteur dont le neuromédiateur est la noradrénaline
pour le système orthosympathique et l'acétylcholine pour le système parasympathique
(récepteur muscarinique).
Le contrôle supra-segmentaire est réalisé par la réticulée, l'hypothalamus,
le système limbique et le cortex pré-frontal.
Le mode de fonctionnement peut être :
- réflexe (afférence - centre - efférence), il correspond aux réflexes de base
- automatique sous contrôle de la réticulée et de l'hypothalamus , assurant
une modulation inconsciente, et capable de réaliser certains programmes

3
- conscient avec un contrôle cortical.
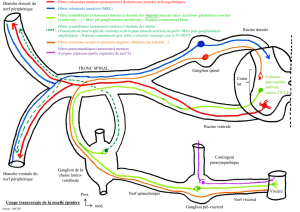
Le système Orthosympathique
Les centres axiaux
La moelle végétative est organisée en colonne longitudinale située au niveau des couches VII de
Rexed
La colonne intermédio médiale s'étend de C1 jusqu'au cône terminal.
Elle est juxta-épendymaire et reçoit les afférences sensitives et nociceptive.
Des inter-neurones segmentaires sont en relation soit avec les moto-neurones alpha soit avec
les colonnes intermédio-latérales. Ils expliquent les arcs réflexes segmentaires à point de
départ sensitif (viscéral ou somatique) et moteur (contracture musculaire réflexe des douleurs
viscérales).
Des inter-neurones connectant deux segments sus et sous jacents.
La colonne intermédio latérale s'étend de C8 à L2.
Elle se divise en une zone antérieure viscéro-motrice et une zone postérieure viscéro-
sensitive
C'est le point de départ des fibres pré-ganglionnaires myélinisées.
Les centres sont successivement :
- le centre cilio spinal de Budge de C8 à T2
- le centre bronchopulmonaire de T3 à T5
- le centre splanchnique abdominopelvien de T6 à L2.
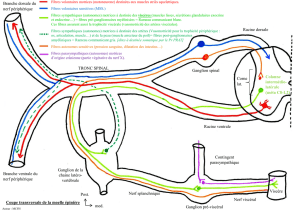
La chaine latérovertébrale :
Les fibres pré-ganglionnaires myélinisées suivent la racine ventrale puis le rameau
communiquant blanc jusqu'au ganglion paravertébral ( chaîne caténaire latéro vertébrale).
Il y a 14 rameaux communiquants blancs correspondant aux 14 étages de D1 à L2 alors qu'il
existe 25 ganglions. Les fibres pré-ganglionnaires remontent ou descendent à differents
niveaux (diffusion et convergence )
Pour le contingent somatique
Les fibres pré-ganglionnaires font relais dans le ganglion paravertébral.
Les fibres post-ganglionnaires suivent le rameau communiquant gris et reviennent jusqu'au
nerf spinal et la disposition métamérique est identique à celle du système nerveux cérébro-
spinal.
Elles assurent l'innervation somatique végétative pour tout le revêtement cutané (vaso-
motricité des vaisseaux sudoripares, pilo-érection).
Un contingent de ces fibres est destiné aux muscles squelettiques , le médiateur terminal est
l'acétylcholine. Lors de la stimulation ortho-sympathique , la libération périphérique
d'Acétydroline est responsable de vaso-dilatation et d'hyper-sudation.
Pour le contingent viscéral

4
Les fibres pré-ganglionnaires traversent les ganglions sans faire relais et se projettent dans
les ganglions pré-vertébraux ou ganglions pré-viscéraux. Elles suivent les artères puis les
plexus péri-artériels.
Au niveau abdomino-pelvien, les fibres pré-ganglionnaires sont les nerfs splanchniques.
On distingue :
- Le nerf grand splanchnique qui naît de T6 à T9 et rejoint le ganglion
coeliaque et mésentérique supérieur
- Le nerf petit splanchnique né de T10, T11 et splanchnique inférieur
né de T12 qui rejoignent les ganglions aortico-rénaux
- Les cinq nerfs splanchniques lombaires réunis en un plexus
hypogastrique supérieur qui rejoint le ganglion du plexus
hypogastrique inférieur
- Les deux premiers nerfs splanchniques lombaires vont aussi vers le
ganglion mésentérique inférieur.
Le système Parasympathique
Les centres crâniens et sacrés
Les Centres para-sympathiques crâniens sont annexés au noyau des nerfs crâniens et les fibres pré-
ganglionnaires suivent le trajet de ces nerfs.
On distingue de haut en bas :
- Le noyau accessoire du nerf oculomoteur ou noyau pupillaire d'Edinger Westphall.
Ces fibres rejoignent par le III le ganglion ciliaire et joue un rôle dans la contraction de
l'iris.
- Le noyau salivaire supérieur en regard du noyau facial, les fibres suivent le nerf
jusqu'au ganglion sous-mandibulaire.
- Le noyau lacrymo muco nasal situé au dessus du noyau salivaire supérieur donne
des fibres qui suivent le nerf facial jusqu'au ganglion ptérygopalatin.
- Le noyau salivaire inférieur en regard du noyau glosso-pharyngien envoie des fibres
jusqu'au ganglion otique et module les sécrétions parotidiennes.
- Le noyau dorsal du X situé dans le plancher du 4ème ventricule envoie des fibres
qui suivent le trajet du pneumogastrique et se distribuent aux viscères du cou, du
thorax et de la plus grande partie de l'abdomen à l'exception du colon et du sigmoïde.
Au niveau sacré le centre est situé dans la colonne ventrale localisée de S2 à S4 . Les fibres pré-
ganglionnaires quittent la moelle par les racines antérieures et suivent les nerfs somatiques sacrés.
Les efferences parasympathiques
Les fibres pré-ganglionnaires longues suivent le trajet des nerfs crâniens jusqu'aux ganglions pré-
viscéraux, à proximité des viscères où ils font relais.

5
Les ganglions
Ces ganglions représentent l'articulation entre le neurone connecteur pré-ganglionnaires et effecteur
post-ganglionnaire.
Ils occupent trois situations :
1° - Les ganglions para-vertébraux de la chaîne caténaire situés de part et d'autre
de la colonne vertébrale allant du crâne au coccyx.
Au niveau cervical, on note deux ou trois ganglions représentés par le ganglion
cervical supérieur en regard de C3, le ganglion cervical moyen inconstant et le
ganglion cervical inférieur ou stellaire en regard de C6.
On note au niveau thoracique 12 ganglions dorsaux, puis 3 - 4 lombaires et enfin 4 ou
5 sacrés. Un ganglion coccygien médian occupe la face antérieure du coccyx.
2° - Les ganglions pré-vertébraux
Ils s'organisent en une chaîne discontinue d'éléments disparates.
Au niveau cervical, ils sont accollés en continuité et forment la partie
antérieure du ganglion cervical supérieur et stellaire.
Au niveau thoracique, ils forment une chaîne co-latérale interne accollée mais
distincte de la chaîne caténaire.
Au niveau abdominal, on distingue les ganglions coeliaques (semi-lunaires),
les ganglions aortico-rénaux, mésentériques supérieurs et mésentériques
inférieurs.
Le plexus solaire réalise de larges mailles qui réunissent entre eux les ganglions
coeliaques et aortico-rénaux, alors que le plexus inter-mésentérique réunit les 2
ganglions du même nom.
Au niveau pelvien, les ganglions pelvipérinéaux sont situés dans les mailles du plexus
hypogastrique inférieur. Ils s'anastomosent avec le ganglion mésentérique inférieur
par le plexus hypogastrique supérieur et les nerfs pré-sacrés.
3° - Les ganglions terminaux pré-viscéraux (ortho et parasympathique).
Ils sont situés à proximité des viscères.
Au niveau céphalique, on distingue le ganglion ciliaire, sphénopalatin, otique
et sous-mandibulaire. Ils sont visibles et topographiquement bien définis.
Au niveau cervical, thoraco-abdominal et pelvien, les ganglions sont petits,
disséminés à la surface des viscères. On distingue le plexus bronchique, le
plexus pharyngien, le plexus cardiaque, vésical, utéro-vaginal.
Au niveau du tube digestif ces ganglions pré-viscéraux sont situés dans
l'épaisseur de la paroi des viscères directement dans la musculeuse.
La médullo-surrénale représente un ganglion terminal dont les éléments cellulaires
sont modifiés dans le sens d'une neurosécrétion adrénergique.
Les voies afférentes du SNV
Les voies afférentes végétatives naissent par des terminaisons libres au niveau des viscères et des
intérocepteurs artériels.
La voie afférente rejoint soit les centres médullaires (ortho-sympathique), soit les centres (para-
sympathique)du tronc cérébral ou de la moelle sacrée.
Les fibres qui rejoignent les centres végétatifs du tronc cérébral, annexées aux nerfs crâniens suivent
le trajet des nerfs crâniens (le glosso-pharyngien, le pneumogastrique).
 6
7
8
9
10
11
12
6
7
8
9
10
11
12
1
/
12
100%