19.11.11 UE 1 Correctionpopulaire

CORPORATION DES ETUDIANTS EN MEDECINE DE PARIS 6
TUTORAT SANTE DE PSA
SAMEDI 19 NOVEMBRE 2011
UE 1 : Atomes, Biomolécules, Génome,
Bioénergétique, Métabolisme
CHIMIE ORGANIQUE – CHIMIE GÉNÉRALE - BIOCHIMIE
CORRECTION
Durée : 1 heure 30 minutes
Documents et calculatrices interdits
Ne peut être vendu ou utilisé dans un but commercial sous peine de poursuite.
Cette correction a été entièrement réalisée par le Tutorat
Ni les professeurs ni la faculté ne pourront être tenus responsables de la validité des
informations qu'elle contient, même en cas d'une éventuelle relecture par un professeur.
Biochimie
14) CDE
15) DE
16) AC
17) D
18) AB
19) BE
20) AD
21) ACE
22) ABC
23) ADE
24) BCD
25) BCE
26) ABE
27) B
28) AC
29) BC
30) ACE
31) ABCD
32) ACE
33) C
34) ABCDE
35) BCD
36) BC
37) B
38) ABC
39) BDE
40) CD
41) C
42) ACD
43) D
44) ABDE
Chimie O
1) ACD
2) CE
3) BC
4) AD
5) D
6) D
Chimie G
7) C
8) DE
9) AD
10) BD
11) ACD
12) BC
13) D

Au programme cette semaine :
Chimie Générale :
o Tout le programme
Chimie Organique :
o Fonctions Divalentes
Biochimie
o Tout le programme
Bon Courage !

CHIMIE ORGANIQUE
QCM 1 et 2
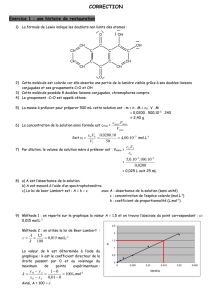
Voici le mécanisme réactionnel qui était à trouver, il s’agissait (attention attention)
d’une imination ! (c’est joli comme mot non ?)
R1O+H++NH2
H
COOH
R2
HOOC N+
OH
R1
H
H
H
R2
C
F
H+
+
H+
D + E
R1O+HR1OH
+
[]
HOOC N
O+
R1
H
H
R2
H
H
HOOC N
O
R1
H
H
R2
H
+
A
B
HOOC N+
R1
H
H
R2
OH2HOOC N
R1
H
R2+
Glucose Acide aminé de l'hémoglobine
1) ACD
A. Vrai, cf diapo 104
B. Faux, le proton s’ajoute à la fonction hydroxyle pour former une molécule d’eau, qui est
un très bon groupe partant
C. Vrai, cf B
D. Vrai, il y a élimination d’eau, comme dans toute imination
E. Faux, cf D. Il ne fallait pas confondre cette réaction avec une acétalisation…

2) CE
A. Faux. L’imination fait toujours intervenir une amine primaire, en effet on se rend compte
que les deux protons liés à l’azote partent, le premier à la deuxième étape, le second à la
sixième (et dernière) étape
B. Faux. Question un peu difficile, ici l’attaque nucléophile vient de l’amine primaire.
Rappelez-vous qu’un dérivé carbonylé est souvent plus apte à se faire attaquer à cause
du carbone lié qui subit les effets inductif et mésomère attracteurs exercés par l’oxygène.
Ce qui entraîne que le carbone tend vers une forme de résonance de type carbocation (cf
qcm 1A) et donc devient un site hautement électrophile, donc très attaquable par un
nucléophile (ici une amine).
C. Vrai, le piège était de ne pas penser à l’acétalisation
D. Faux, c’est une amine primaire qui attaque, pas un alcool
E. Vrai, l’imination est une déshydratation
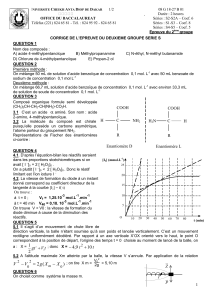
QCM 3 à 6
Mécanisme
- Cétolisation en milieu basique :
1ère étape : Déprotonation de la cétone
CH3O
H+BCH3
OBH
+
O
CH3
CH2
1 a
2
b
2ème étape : Attaque nucléophile de l’énolate sur l’aldéhyde
CH3
OH
O
+
OO
234
3ème étape : Réaction acido-basique → Obtention d’un cétol (5)

OO
4+BH
b
OH O
+B
5a
Crotonisation :
4ème étape : Réaction acido-basique
OH O
H+B
5a
OH O
+BH
b
Elimination intramoléculaire du groupe hydroxyle
OH O
O
+OH
6
5ème étape (non détaillée dans le sujet) : Régénération de la base
BH
b
OH +B
a+H2O
3) BC
A. Faux : Zéro carbone asymétrique sur cette molécule !
B. Vrai : a correspond au catalyseur de la réaction, il s’agit d(une base forte (milieu basique).
En effet, OH- fera bien l’affaire !
C. Vrai : cf mécanisme
D. Faux : il s’agit d’une réaction acido-basique (cf mécanisme)
E. Faux : L’attaque nucléophile a lieu dans la 2ème étape (2ème double flèche). Deux réactifs
et un seul produit, cela aurait dû vous dire quelque chose ;)
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
6
7
8
9
10
11
12
13
14
15
16
17
18
19
1
/
19
100%