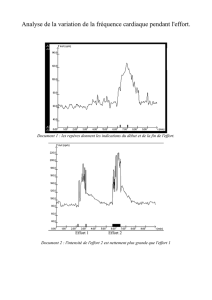
III. Physiologie rénale - Formation de l`urine

Physiologie animale Pr. Tobias Hevor Semestre 5
1
Professeur Tobias Hevor
Directeur du Laboratoire de Neurobiologie
Université d’Orléans. B.P. 6759
45067 Orléans CEDEX 2
L3 : Les grandes
fonctions
Milieu intérieur
Physiologie cardiaque
Physiologie vasculaire
Respiration
Excrétion
Digestion
Nutrition

Physiologie animale Pr. Tobias Hevor Semestre 5
2
Sommaire
Professeur Tobias Hevor ......................................................................................................................... 1
Milieu intérieur ........................................................................................................................................ 1
Sommaire ................................................................................................................................................ 2
Chapitre 0 : Enthalpie et Entropie ........................................................................................................... 7
Chapitre 1 : Le milieu intérieur ................................................................................................................ 8
I. Les compartiments liquidiens de l'organisme ................................................................................. 8
A. Un compartiment ........................................................................................................................ 8
B. Deux compartiments ................................................................................................................... 9
C. Compartiments multiples ............................................................................................................ 9
D. Composition des liquides et mouvements liquidiens ............................................................... 10
II. Le Sang ........................................................................................................................................... 12
A. Propriétés physiques et mesure du volume sanguin ................................................................ 12
B. Le plasma sanguin et sa composition ........................................................................................ 13
1. La composition du plasma ..................................................................................................... 13
2. Les substances protéiques .................................................................................................... 13
3. Composés non-protéiques .................................................................................................... 13
4. Système tampon du sang ...................................................................................................... 14
5. Le système des protéines ...................................................................................................... 14
6. Le système imidazole ............................................................................................................ 15
7. Le système effet Bohr ............................................................................................................ 15
8. Le système de l’acide carbonique ......................................................................................... 15
C. Origine des éléments figurés ..................................................................................................... 15
1. Les érythrocytes .................................................................................................................... 15
2. Les leucocytes ........................................................................................................................ 15
3. Les thrombocytes .................................................................................................................. 16
III. Les éléments figurés .................................................................................................................. 17
A. Les globules rouges (ou hématies ou érythrocytes).................................................................. 17
1. Caractéristiques ..................................................................................................................... 17
2. Régulation de la production des hématies ............................................................................ 17
3. Eléments constitutifs des globules rouges ............................................................................ 18
4. Rôles des pigments du sang .................................................................................................. 21

Physiologie animale Pr. Tobias Hevor Semestre 5
3
B. Les plaquettes sanguines et la coagulation du sang ................................................................. 21
1. Caractéristiques ..................................................................................................................... 21
2. Hémostase ............................................................................................................................. 21
3. Les leucocytes ........................................................................................................................ 23
4. Les groupes sanguins ............................................................................................................. 26
Chapitre 2 : La physiologie cardiaque ................................................................................................... 28
I. Généralités sur l’appareil cardio-vasculaire .................................................................................. 28
II. Rappel de l’anatomie cardiaque .................................................................................................... 28
A. Morphologie et structure .......................................................................................................... 28
B. Histologie ................................................................................................................................... 29
III. Fonctionnement global du cœur ............................................................................................... 29
A. Activité électrique ..................................................................................................................... 29
B. Activité mécanique et sa genèse ............................................................................................... 30
IV. Propriétés du cœur .................................................................................................................... 31
A. Automatisme cardiaque et rythmicité ...................................................................................... 31
1. Origine de l’automatisme ...................................................................................................... 31
2. Mécanisme de la rythmicité .................................................................................................. 33
3. Régulation de la rythmicité ................................................................................................... 33
4. Potentiel d’action des cellules ventriculaires ........................................................................ 34
B. Conductibilité ............................................................................................................................ 35
1. Propagation de l’influx .......................................................................................................... 35
2. Arrêt de la propagation ......................................................................................................... 35
V. Régulation de l’activité cardiaque ................................................................................................. 36
A. Régulation de la fréquence ....................................................................................................... 36
1. Innervation cardiaque ........................................................................................................... 36
2. Régulation.............................................................................................................................. 37
3. Rôle des hormones ................................................................................................................ 37
B. Régulation du volume de l’ondée systolique ............................................................................ 37
VI. Electrocardiographie ................................................................................................................. 37
Chapitre 3 : Physiologie vasculaire ........................................................................................................ 39
I. Anatomie des vaisseaux ................................................................................................................ 39
A. Structure de la paroi vasculaire ................................................................................................. 39
B. Propriétés de la paroi vasculaire ............................................................................................... 39
II. Hémodynamique ........................................................................................................................... 40

Physiologie animale Pr. Tobias Hevor Semestre 5
4
III. Circulation artérielle .................................................................................................................. 41
A. Propriétés .................................................................................................................................. 41
B. Mesure de la pression artérielle ................................................................................................ 41
1. Méthode directe .................................................................................................................... 41
2. Méthode indirecte ................................................................................................................. 42
C. Le pouls ...................................................................................................................................... 43
D. Régulation de la pression artérielle ........................................................................................... 43
1. Facteurs régulant la pression artérielle ................................................................................. 43
2. Mécanisme nerveux de la régulation de la pression artérielle ............................................. 44
E. Circulation capillaire – Equilibre de Starling .............................................................................. 46
F. Circulation veineuse .................................................................................................................. 48
Chapitre 4 : Physiologie de la respiration .............................................................................................. 50
I. Lois physiques ................................................................................................................................ 50
II. Mécanique ventilatoire ................................................................................................................. 50
A. Structure de l’appareil respiratoire ........................................................................................... 50
B. Modifications de volume ........................................................................................................... 54
C. Modification de pression ........................................................................................................... 55
D. Volumes et capacités respiratoires ........................................................................................... 55
E. Composition de l’air atmosphérique et de l’air alvéolaire ........................................................ 56
III. Transport des gaz ...................................................................................................................... 56
A. Transport de l’oxygène .............................................................................................................. 56
B. Transport du CO2 ....................................................................................................................... 59
IV. Régulation de la ventilation....................................................................................................... 60
A. Rythmicité ................................................................................................................................. 60
B. Facteurs modifiant l’activité des centres. ................................................................................. 62
Chapitre 5 : Excrétion ............................................................................................................................ 64
I. Structure macroscopique .............................................................................................................. 64
II. Le Néphron .................................................................................................................................... 67
A. Morphologie .............................................................................................................................. 67
B. Disposition spaciale ................................................................................................................... 69
C. L’appareil justaglomérulaire ...................................................................................................... 70
III. Physiologie rénale - Formation de l’urine ................................................................................. 70
A. Filtration glomérulaire ............................................................................................................... 71
B. Formation de l’urine définitive .................................................................................................. 72

Physiologie animale Pr. Tobias Hevor Semestre 5
5
1. Réabsorption et sécrétion tubulaire ..................................................................................... 72
2. Mécanisme de concentration de l’urine ............................................................................... 73
IV. Régulation des paramètres du milieu intérieur ........................................................................ 75
I. La cavité buccale et le pharynx...................................................................................................... 77
A. Anatomie de la cavité buccale ................................................................................................... 77
B. Le pharynx et la déglutition ....................................................................................................... 79
C. Les glandes salivaires ................................................................................................................. 79
1. Organisation .......................................................................................................................... 79
2. Activité des glandes salivaires ............................................................................................... 81
D. Absorption ................................................................................................................................. 82
II. Œsophage ...................................................................................................................................... 82
III. L’estomac .................................................................................................................................. 84
A. Aspect morphologique, histologique et cytologique ................................................................ 84
B. Vascularisation et innervation ................................................................................................... 86
C. Sécrétion gastrique.................................................................................................................... 86
1. Le suc gastrique ..................................................................................................................... 86
2. Inhibition de la sécrétine ....................................................................................................... 86
3. Facteurs intervenant dans la sécrétion ................................................................................. 86
4. Aspect enzymatique de la sécrétion ..................................................................................... 86
5. Absorption – Vidange ............................................................................................................ 86
IV. L’intestin .................................................................................................................................... 87
A. Aspects anatomiques ................................................................................................................ 87
B. Le pancréas exocrine ................................................................................................................. 88
1. Aspects anatomiques ............................................................................................................ 88
2. Sécrétions pancréatiques ...................................................................................................... 88
C. La sécrétion biliaire ................................................................................................................... 89
1. Aspect anatomique du foie ................................................................................................... 89
2. La bile..................................................................................................................................... 91
D. La digestion intestinale .............................................................................................................. 92
1. Les glucides ............................................................................................................................ 92
2. Les protides ........................................................................................................................... 92
3. Les lipides .............................................................................................................................. 92
4. Mouvements d’eau ............................................................................................................... 93
5. Sels minéraux ........................................................................................................................ 93
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
1
/
95
100%