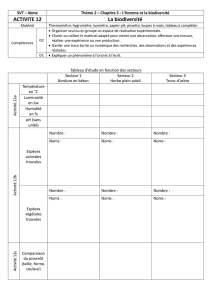
la diversité écologique d`un écosystème dans un paysage agricole

DANIEL LACHANCE
LA DIVERSITÉ ÉCOLOGIQUE D’UN ÉCOSYSTÈME DANS
UN PAYSAGE AGRICOLE :
LE CAS DES TOURBIÈRES DU BAS-SAINT-LAURENT
(QUÉBEC)
Thèse
présentée
à la Faculté des études supérieures
de l’Université Laval
pour l’obtention
du grade de Philosophiæ Doctor (Ph.D.)
École supérieure d’aménagement du territoire et de développement régional
FACULTÉ D’AMÉNAGEMENT, D’ARCHITECTURE ET DES ARTS VISUELS
UNIVERSITÉ LAVAL
QUÉBEC
FÉVRIER 2005
© Daniel Lachance, 2005

II
Résumé
Cette thèse est une étude en écologie appliquée comportant une perspective spécifique à
l’aménagement du territoire. L’objectif principal est de comprendre comment l’isolement d’un
écosystème au sein d’un territoire perturbé par les activités humaines modifie sa diversité
écologique, tant sur le plan végétal qu’animal. Dans un premier temps, des analyses
multivariées sont utilisées afin de départager l’influence des activités anthropiques et des
variables abiotiques sur la végétation des tourbières ombrotrophes de la région de Rivière-du-
Loup – L’Isle-Verte, dans le Bas-Saint-Laurent (Québec, Canada). Ces analyses montrent
l’existence d’une gradient allant des grands sites ouverts pas ou très peu perturbés aux petits
sites boisés où la végétation est clairement influencée par les activités humaines. Les résultats
soulignent l’influence manifeste des activités anthropiques, et plus particulièrement de
l’agriculture, sur les assemblages végétaux. Dans un deuxième temps, l’impact du boisement
des tourbières, une conséquence des perturbations anthropiques, est évalué chez les plantes et
les oiseaux. Les résultats soulignent l’impact négatif du boisement sur les plantes
caractéristiques des écosystèmes tourbeux et sur les espèces d’oiseaux nichant de préférence
dans les tourbières, telles le bruant de Lincoln et la paruline masquée. Malgré tout, les
tourbières du Bas-Saint-Laurent demeurent des îlots de végétation boréale dans un paysage
constitué de forêts décidues et de champs agricoles. Enfin, les résultats obtenus sont placés
dans leur contexte régional et la pertinence d’un projet de conservation est abordée. Peu
d’espèces de plantes et d’oiseaux sont confinées aux tourbières dans le Bas-Saint-Laurent.
Cependant, le caractère unique de ces îlots tourbeux, leur rôle de refuge écologique et la rareté
des aires protégées dans le Bas-Saint-Laurent justifient amplement l’attribution d’un statut de
conservation à la tourbière de Bois-des-Bel, la plus intègre des tourbières de la région.
Globalement, la thèse illustre de façon claire l’importance d’une base de données très détaillée
sur les organismes vivants pour cerner les impacts – parfois subtils mais pas pour autant
négligeables – des activités humaines sur la diversité écologique. La thèse met également en
relief la nécessité de considérer le contexte du paysage régional afin de bien comprendre les
répercussions des activités humaines perturbatrices sur les écosystèmes.

III
Avant-propos
Le corps de la thèse comporte trois chapitres, dont deux rédigés en anglais (Chapitres II et
III) et ayant la forme d’articles scientifiques. Le Chapitre II, portant sur les facteurs qui
influencent la végétation des écosystèmes tourbeux de la région de Rivière-du-Loup – L’Isle-
Verte, a été publié en 2004 dans la revue Applied Vegetation Science. Le Chapitre III, portant
sur l’impact du boisement des tourbières sur les espèces de plantes et d’oiseaux, a été accepté
pour publication dans la revue Écoscience et paraîtra en 2005. Mon directeur de recherche,
M. Claude Lavoie, professeur à l’École supérieure d’aménagement du territoire et de
développement régional, est co-auteur des Chapitres II et III. Mon codirecteur, M. André
Desrochers, professeur au Département des sciences du bois et de la forêt, est pour sa part co-
auteur du Chapitre III. Néanmoins, j’atteste être le principal responsable de l’élaboration des
travaux de recherche, du traitement et de l’analyse des données et de la rédaction des
manuscrits.
La stylistique de l’ensemble des articles, le nom des sites et leur abréviation ont été
modifiés afin d’harmoniser l’ensemble du contenu de la thèse. Afin de faciliter la lecture
individuelle des différents chapitres, il a été décidé de conserver les articles dans leur
intégralité. Il existe donc une certaine redondance entre les chapitres, notamment dans les
sections décrivant la méthodologie.

IV
Remerciements
Cette thèse est sans contredit le fruit de mes efforts, mais sa réalisation fut en bonne partie
possible grâce à l’entière et gracieuse collaboration de plusieurs personnes. En tout premier
lieu, je tiens à remercier mon directeur de thèse, M. Claude Lavoie, pour le support
pédagogique et technique qu’il a su m’apporter. Je lui suis également reconnaissant pour la
confiance dont il a fait preuve en m’offrant de poursuivre un doctorat avec lui. Mentionnons
également la participation de mon codirecteur, M. André Desrochers, qui m’a prodigué de
judicieux conseils à chacune de nos rencontres. Mes remerciements vont également aux autres
membres de mon comité de supervision, soit MM. Marius Thériault, André Bouchard et Jean-
Pierre Savard et à Mme Monique Poulin pour l’intérêt qu’ils ont porté à mes recherches.
Je remercie particulièrement mes assistants de terrain, soit Vicky Albert, Magali Crevier,
Yvon Jodoin, Laurence Letarte et Claudia Zimmermann. Ce fut un honneur (et un plaisir!) de
vivre à leur côté pendant deux étés. La qualité de mes données découle en grande partie de
leur dévouement et de leur rigueur. Mes remerciements vont également à MM. Robert
Gauthier et Claude Roy et à Mme Michelle Garneau de l’Herbier-Louis-Marie, pour leur
support et leur coopération dans l’identification d’un nombre impressionnant de sphaignes,
mousses, hépatiques, lichens et autres végétaux. Je suis particulièrement reconnaissant envers
M. Marc Girard, pour l’aide précieuse apportée dans mes analyses multivariées et envers
M. Bruno Drolet, pour son aide en matière d’échantillonnage de la faune aviaire.
J’aimerais également souligner la chaleureuse participation de Mme Stéphanie Pellerin, non
seulement pour le prêt gracieux de ses données historiques, mais également pour l’aide et les
conseils précieux apportés à chaque étape de la réalisation de cette thèse. Après moi,
Stéphanie est certainement la personne qui a lu le plus de versions préliminaires et de
manuscrits issus de mes travaux. Ce doctorat n’aurait pas été le même sans son support.
Je tiens à remercier tous les membres actuels et passés du Laboratoire d’écologie historique
(LEH), du Groupe de recherche en écologie des tourbières (GRET) et du Centre de recherche
en aménagement et développement (CRAD) pour leur support tant académique que technique
et pour le cadre de recherche stimulant dans lequel ils m’ont permis de travailler. Parmi ceux-

V
ci, je tiens particulièrement à exprimer ma gratitude envers Fanny Delisle, Pierre Lemieux,
Kathleen Marcoux et François Théberge pour les services rendus et les nombreuses
discussions.
J’adresse des remerciements particuliers à ma famille et à mes amis qui m’ont encouragé
tout au long de mes études. Parmi eux je m’en voudrais de ne pas souligner l’aide précieuse de
ma conjointe, Adeline, dont l’enthousiasme, la curiosité et le support furent d’une aide
inestimable.
Enfin, cette recherche a été possible grâce au soutien financier du Centre de recherche en
aménagement et développement, de la Fondation de l’Université Laval, de la Fondation
Marie-Victorin, du Conseil de recherches en sciences naturelles et en génie du Canada et de
l’industrie canadienne de la tourbe.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
1
/
117
100%