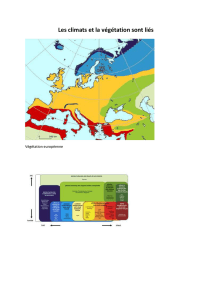
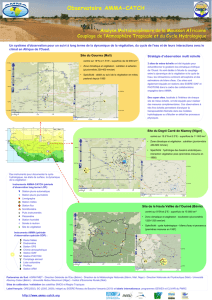
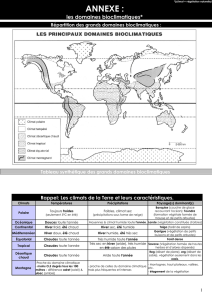
Dynamique de la biodiversité végétale dans les paysages d

UNIVERSITE DE TOULOUSE II – LE MIRAIL
UFR SCIENCES – ECONOMIE - SOCIETE
DEPARTEMENT GEOGRAPHIE – AMENAGEMENT
2004 N° attribué par la bibliothèque
THESE pour obtenir le grade de
Docteur de l’Université de Toulouse II – Le Mirail
Discipline : Géographie – Aménagement
Présentée et soutenue publiquement par
Brigitte SCHWAL
le 5 juillet 2004
Dynamique de la biodiversité végétale dans les
paysages d’agriculture intensive
Utilisation du sol, composition floristique, végétation et structures paysagères du
Lauragais haut-garonnais (Sud-Ouest, France), XIXe-XXe siècles
Dynamics of plant biodiversity in intensive farming landscapes
Land-use, floristic composition, vegetation and landscape’s pattern of Lauragais
haut-garonnais (South-west of France), XIXth-XXth centuries
Sous la direction de
M. Jacques HUBSCHMAN
M. Thierry GAUQUELIN
Devant la commission d’examen composée de :
J. BAUDRY (Rapporteur) Directeur de recherche à l’INRA-SAD Armorique
T. GAUQUELIN (Directeur de Thèse) Professeur à l’Université Paul Sabatier Toulouse III
C. GERS (Examinateur) Chargé de recherche CNRS
J. HUBSCHMAN (Directeur de Thèse) Professeur à l’Université Toulouse II – Le Mirail
G. LARGIER (Examinateur) Directeur du CBN Midi-Pyrénées
J-P. METAILIE (Examinateur) Directeur de recherche CNRS
J. MORET (Rapporteur) Professeur du Muséum National d’Histoire Naturelle
GEODE Ecole Doctorale LADYBIO
UMR5602 UT2-CNRS Temps-Espaces-Sociétés-Cultures UMR5172 UT3-CNRS

2
Remerciements
Je tiens à adresser mes remerciements aux nombreuses personnes qui ont apporté à leur
niveau une contribution à ce travail.
En premier lieu, je témoigne toute ma gratitude envers mes directeurs de thèse, Mrs T.
Gauquelin (Professeur à l’Université Paul Sabatier à Toulouse) et J. Hubshman (Professeur à
l’Université de Toulouse-Le Mirail), de m’avoir accorder leur confiance, de m’avoir permis
d’exécuter ce travail dans les meilleures conditions, de leur disponibilité à chaque fois que je
les ai sollicité, ainsi que de leurs conseils avisés.
Mes remerciements vont également à M. C. Gers, Chargé de recherche CNRS, que j’ai eu
l’occasion de rencontrer pour la première fois lors d’un stage à l’ancien Laboratoire
d’Ecologie Terrestre et lors duquel il me présenta un inventaire floristique datant de 1885,
document qui devint le point de départ de ma recherche. Je le remercie grandement de ses
encouragements, du temps consacré à me conseiller vers certaines directives de recherche et
de son soutien plus qu’une fois heureux. Mon immersion dans le Lauragais haut-garonnais qui
a commencé en DEA, s’est poursuivi en thèse au laboratoire GEODE, grâce à l’obtention
d’une allocation de recherche. Ma profonde reconnaissance va à M. J-P Métailié, Directeur du
laboratoire GEODE, pour sa bienveillance et ses conseils avisés. Je remercie ici l’ensemble
des membres du laboratoire GEODE qui ont toujours été à mes côtés pour me soutenir et
m’aider dès qu’ils le pouvaient. Je tiens à remercier en particulier M. M. Paegelow, Maître de
conférence à l’Université de Toulouse-Le mirail, de son soutien et de sa relecture critique.
Je remercie tout particulièrement Ms P. Le Caro, Ingénieur d’étude CNRS et L. Belhacène,
Responsable de l’association Isatis, de leur aide précieuse lors du grand plongeon dans l’étude
de la flore et notamment des herborisations sur le terrain et des déterminations, ainsi que de
leur disponibilité à chaque fois que j’ai eu à solliciter leurs connaissances de naturalistes
avertis.
Ma formation aux traitements statistiques a été instruite par M. A. Fabre, Ingénieur d’étude
CNRS, à qui je rends hommage. Je remercie plus généralement l’équipe du LADYBIO qui
m’a aidé et soutenu par ses compétences, notamment M. S. Maumont du temps qu’il a bien
voulu consacré au partage de ses connaissances botaniques. Le CBN Midi-Pyrénées m’a
permis de constituer une base de données à partir de ses références. Je remercie son directeur
M. G. Largier et l’ensemble de l’équipe qui m’ont aidé à un moment ou à un autre dans mon
travail. Mes remerciements vont également à l’équipe DYNAFOR de l’INRA de Toulouse,
qui a fait avancer mes réflexions.
Merci à Monsieur Le Maire de Nailloux, Georges Méric, à son conseiller municipal, M. A.
Marty ainsi qu’aux responsables du Centre Culturel du Lauragais, Mme A. Nivet-Sanger, M.
M. Dutech et M. J. Odol, pour l’intérêt qu’ils ont accordé à mon travail.
Je tiens également à exprimer ma profonde gratitude à Ms J. Baudry (Directeur de recherche
INRA) et J. Moret (Professeur au MNHN) qui m’ont fait l’honneur d’être rapporteurs de cette
thèse.
Enfin, l’aboutissement de ce travail est aussi le fruit du soutien et parfois de la patience de
mes proches. Merci à Benjamin, Gisèle, Marianne et Bernard.

3
Table des matières
Remerciements.......................................................................................................................2
Liste des figures .....................................................................................................................8
Liste des tableaux.................................................................................................................10
Liste des photographies........................................................................................................11
Abréviations spécifiques à la thèse ......................................................................................12
Introduction..........................................................................................................................13
1 - Contexte de l’étude : la biodiversité végétale dans les paysages d’agriculture intensive ............ 13
2 - Hypothèses et objectifs de l’étude : impact des pratiques agricoles et de leur évolution sur la
biodiversité, au cours des deux derniers siècles........................................................................... 14
3 - Méthodologie de recherche et organisation du mémoire............................................................. 17
Chapitre 1 La biodiversité, un concept émergent et son expression
dans les paysages d’agriculture intensive ................................................21
I - Le concept émergent de biodiversité...............................................................22
1 - Origines du concept de biodiversité............................................................................................. 22
2 - Définition du concept de biodiversité.......................................................................................... 23
3 - Textes juridiques relatifs à la protection de la biodiversité.......................................................... 23
4 - Niveaux d’organisation de la biodiversité ................................................................................... 24
4. 1 - Niveau intraspécifique......................................................................................................... 25
4. 2 - Niveau spécifique ................................................................................................................ 26
4. 3 - Niveau interspécifique......................................................................................................... 26
4. 4 - Niveau supra-spécifique ...................................................................................................... 26
5 - Importance de la biodiversité végétale ........................................................................................ 26
5. 1 - Préoccupations et menaces pesant sur la biodiversité végétale............................................ 27
5. 2 - Valeur des composantes de la biodiversité végétale............................................................ 28
5. 2. 1 - Valeur comme fonction au sein de l’écosystème........................................................................ 28
5. 2. 2 - Valeur pour l’économie et la santé............................................................................................ 29
5. 2. 3 - Valeurs de caractère éthique et culturel.................................................................................... 29
5. 3 - Évaluer la biodiversité : Considérations qualitatives et quantitatives.................................. 30
6 - Facteurs influençant la biodiversité végétale et la répartition de la végétation............................ 32
6. 1 - Facteurs mésologiques......................................................................................................... 32
6. 2 - Facteurs anthropiques.......................................................................................................... 32
II - L’expression de la biodiversité végétale dans les paysages d’agriculture
intensive .............................................................................................................33
1 - Comment l’expression de la biodiversité végétale est-elle possible dans des paysages fortement
anthropisés ?................................................................................................................................ 34
1. 1 - L’Ecologie du paysage, ou l’étude des effets des composantes de la mosaïque paysagère sur
la biodiversité végétale.................................................................................................................. 35
1. 1. 1 - Les perturbations, sources d’hétérogénéité spatio-temporelle.................................................. 36
1. 1. 2 - La fragmentation....................................................................................................................... 39
1. 1. 3 - La connectivité........................................................................................................................... 40
1. 2 - Rôle de la variabilité génétique et de la banque de graine, dans la colonisation d’habitats. 41
2 - Conséquences des pratiques agricoles sur la biodiversité végétale.............................................. 42
2. 1 - Impact négatif de la modernisation des paysages agricoles sur la biodiversité végétale..... 42
2. 1. 1 - La dégradation des habitats ...................................................................................................... 43
2. 1. 2 - La réduction du nombre d’espèces utilisées.............................................................................. 43
2. 1. 3 - La disparition de la vie sauvage................................................................................................ 43

4
2. 2 - Les pratiques agricoles peuvent être initiatrices de biodiversité.......................................... 44
III - Réflexion sur l’importance de la valeur écologique des espèces dans une
optique conservatoire........................................................................................45
1 - Abondance des espèces dans le milieu ........................................................................................ 45
2 - Valeur patrimoniale des espèces.................................................................................................. 46
3 - Origine biogéographique des espèces.......................................................................................... 47
Chapitre 2 Le territoire d’étude : le Lauragais haut-garonnais, Pays
d’agriculture intensive...............................................................................49
I - Limites du Lauragais haut-garonnais.............................................................50
II - Identité paysagère du Lauragais haut-garonnais ..........................................53
1 - Géomorphologie.......................................................................................................................... 53
2 - Hydrographie............................................................................................................................... 54
3 - Grands traits du climat................................................................................................................. 54
4 - Pédologie..................................................................................................................................... 55
5 - Végétation.................................................................................................................................... 55
6 - Économie..................................................................................................................................... 56
7 - Population.................................................................................................................................... 56
III - Les sites d’étude, représentatifs du Lauragais haut-garonnais....................57
1 - Le site de Nailloux....................................................................................................................... 57
2 - Le site de Préserville.................................................................................................................... 58
IV - Éléments pour une histoire de l’agriculture lauragaise ................................58
1 - Du Néolithique au Moyen-Âge : les grands traits de la mise en place des paysages agraires..... 58
2 - De la fin de l’Antiquité à la fin du Moyen-Âge : continuités et restructurations du paysage rural
..................................................................................................................................................... 60
3 - De la fin du XVe au XVIIe siècle, l’âge d’or du pastel............................................................... 61
4 - Du XVIIIe siècle au début du XXe siècle, la diversification des cultures et les progrès des
méthodes...................................................................................................................................... 62
4. 1 - Le XVIIIe siècle, des systèmes agricoles traditionnels........................................................ 63
4. 2 - Au XIXe siècle, les mutations des systèmes agricoles traditionnels.................................... 64
5 - Au XXe siècle, la modernisation de l’agriculture lauragaise....................................................... 66
Chapitre 3 Organisation spatiale de la végétation en réponse à
l’artificialisation du paysage .....................................................................69
I - Acquisition des données spatiales....................................................................70
1 - Intérêt et principe d’un SIG......................................................................................................... 71
1. 1 - Stockage de l’information en mode vecteur ........................................................................ 71
1. 2 - Stockage de l’information en mode raster........................................................................... 72
2 - Inventaire des documents de référence........................................................................................ 72
2. 1 - Échelles d’observation......................................................................................................... 73
2. 2 - Nécessité d’un état de référence .......................................................................................... 73
2. 3 - Documents retenus............................................................................................................... 73
2. 3. 1 - Les documents numériques........................................................................................................ 73
2. 3. 2 - Les documents cartographiques ................................................................................................ 73
2. 3. 3 - Les documents photographiques................................................................................................ 74
3 - Typologie de l’occupation du sol des formations végétales ........................................................ 76
3. 1 - Formations végétales artificialisées..................................................................................... 78
3. 2 - Formations à végétation spontanée...................................................................................... 79
4 - Définition des attributs spatiaux des unités paysagères............................................................... 82
II - Analyse de l’organisation spatiale des formations végétales : une
hétérogénéité des paysages en relation avec les pratiques agricoles ............84

5
1 - Organisation actuelle des formations végétales : analyse synchronique...................................... 84
1. 1 - Étude synchronique des attributs spatiaux........................................................................... 86
1. 1. 1 - Structures du paysage................................................................................................................ 86
1. 1. 2 - Fragmentation des taches boisées............................................................................................. 90
1. 2 - L’hétérogénéité spatiale des paysages influence leur capacité d’accueil............................. 92
2 - Évolution de l’organisation des formations végétales : analyse diachronique............................. 95
2. 1 - Un état de référence du paysage, 1834. Des paysages différemment organisés pour les deux
sites 95
2. 2 - Étude diachronique des attributs spatiaux............................................................................ 98
2. 2. 1 - Structure du paysage................................................................................................................. 98
2. 2. 2 - Fragmentation des taches boisées........................................................................................... 104
2. 3 - Étude des grands traits d’évolution du paysage................................................................. 105
2. 4 - Synthèse : une hétérogénéité temporelle des paysages en relation avec des pratiques
agricoles évolutives..................................................................................................................... 107
III - Les enseignements tirés de l’organisation spatiale de la végétation...........114
1 - L’hétérogénéité du paysage en réponse à son aménagement..................................................... 115
2 - Fragmentation des taches boisées et effet-lisière....................................................................... 116
3 - Les activités humaines, facteurs favorables et / ou préjudiciables aux formations végétales.... 116
4 - Notion de supplémentation versus complémentation ................................................................ 117
5 - Discussion sur la méthode......................................................................................................... 117
Chapitre 4 Variation de la flore et de la végétation en réponse à
l’artificialisation du paysage : composition et diversité floristiques...119
I - Les listes floristiques, documents de base de l’étude de la flore.................121
1 - La flore d’une région, élément d’évaluation de la biodiversité.................................................. 121
1. 1 - Connaissance de la flore actuelle : l’inventaire des taxons................................................ 121
1. 1. 1 - Mise en place des relevés de végétation.................................................................................. 122
1. 1. 2 - Réalisation des relevés de végétation...................................................................................... 127
1. 1. 3 - Limites de la méthode par relevés phytosociologiques............................................................ 129
1. 2 - Connaissance de la flore passée : l’inventaire floristique communal ................................ 129
1. 3 - Liste floristique globale..................................................................................................... 131
1. 4 - Indications bibliographiques : des sources d’information supplémentaires....................... 131
1. 4. 1 - Les documents floristiques anciens.......................................................................................... 131
1. 4. 2 - Les documents floristiques actuels .......................................................................................... 133
1. 5 - Harmonisation nomenclaturale.......................................................................................... 134
2 - Les caractéristiques descriptives de la flore et de la végétation................................................. 134
2. 1 - Les indices de biodiversité, caractéristiques de la flore..................................................... 134
2. 2 - Les données de l’autécologie, descripteurs de la végétation.............................................. 135
II - Mise en évidence de la variation spatio-temporelle de la flore : estimation de
la tendance évolutive des espèces...................................................................138
1 - Aspects actuels de la flore et de la végétation lauragaises......................................................... 138
1. 1 - Caractérisation de la flore.................................................................................................. 139
1. 1. 1 - Richesse spécifique.................................................................................................................. 139
1. 1. 2 - Diversité floristique................................................................................................................. 141
1. 1. 3 - Diversité supraspécifique ........................................................................................................ 141
1. 1. 4 - Richesse patrimoniale.............................................................................................................. 144
1. 1. 5 - Fréquence des espèces............................................................................................................. 146
1. 1. 6 - Synthèse sur la caractérisation de la flore............................................................................... 148
1. 2 - Description de la végétation .............................................................................................. 149
1. 2. 1 - Liste floristique globale........................................................................................................... 149
1. 2. 2 - Liste floristique pour chaque site............................................................................................. 152
1. 2. 3 - Synthèse : des végétations disparates pour les deux sites........................................................ 156
1. 3 - Physionomie de la végétation et identification des formations végétales.......................... 157
1. 4 - Des nuances de la flore actuelle, sous l’influence de schémas d’organisations spatiales
différentes.................................................................................................................................... 159
2 - Variations de la flore et de la végétation lauragaises, depuis 1885............................................ 160
2. 1 - Exploitation de la Flore d’Haulier, Nailloux, 1885............................................................ 160
2. 1. 1 - Caractérisation de la flore....................................................................................................... 161
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
231
232
233
234
235
236
237
238
239
240
241
242
243
244
245
246
247
248
249
250
251
252
253
254
255
256
257
258
259
260
261
262
263
264
265
266
267
268
269
270
271
272
273
274
275
276
277
278
279
280
281
282
283
284
285
286
287
288
289
290
291
292
293
294
295
296
297
298
299
300
301
302
303
304
305
306
307
308
309
310
311
312
313
314
315
316
317
318
319
320
321
322
323
324
325
326
327
328
329
330
331
332
333
334
335
336
337
338
339
340
341
342
343
344
345
346
347
348
349
350
351
352
353
354
355
356
357
358
359
360
361
362
363
364
365
366
367
368
369
370
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
231
232
233
234
235
236
237
238
239
240
241
242
243
244
245
246
247
248
249
250
251
252
253
254
255
256
257
258
259
260
261
262
263
264
265
266
267
268
269
270
271
272
273
274
275
276
277
278
279
280
281
282
283
284
285
286
287
288
289
290
291
292
293
294
295
296
297
298
299
300
301
302
303
304
305
306
307
308
309
310
311
312
313
314
315
316
317
318
319
320
321
322
323
324
325
326
327
328
329
330
331
332
333
334
335
336
337
338
339
340
341
342
343
344
345
346
347
348
349
350
351
352
353
354
355
356
357
358
359
360
361
362
363
364
365
366
367
368
369
370
1
/
370
100%