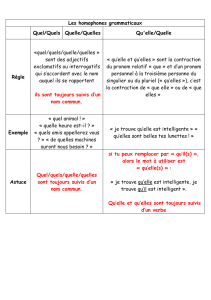
sur le programme de surveillance et plan d

Directive Cadre Stratégie pour le Milieu Marin
Programme de surveillance et
plan d’acquisition de connaissances
Propositions scientifiques et techniques (chantier 2)
Thématique 4 : « Tous les éléments constituant le réseau trophique marin, dans la
mesure où ils sont connus, sont présents en abondance et diversité
normales et à des niveaux pouvant garantir l’abondance des espèces
à long terme et le maintien total de leurs capacités reproductives. »
Rapport de synthèse de l’exercice 2012
Version 2 du 7 juillet 2013
Pilote thématique 4 : CNRS-INEE
Contributions à la rédaction (par ordre alphabétique): Grégory BEAUGRAND, Xavier
FIZZALA, Benoit MIALET, Jean-Pierre FERAL, François LE LOC'H, Nathalie NIQUIL,
Florent RENAUD, Isabelle ROMBOUTS.

Remerciements
Nous tenons à remercier l’ensemble des participants aux ateliers et relecteurs ayant fortement
contribué à l’amélioration de ce rapport.
Atelier « Benthos » : Sophie BEAUVAIS, Antoine CARLIER, Jacques GRALL, Céline
LABRUNE, Sandrine LAURAND, François Le LOC’H, Pascal RIERA, Pierre RICHARD,
Gauthier SCHAAL, Eric THIEBAUT, Céline VIGNOT.
Atelier « Plancton » : Felipe ARTIGAS, Pascal CLAQUIN, Valérie DAVID, Alain
LEFEBVRE, Dominique LEFEVRE, Fabien LOMBARD, Behzad MOSTAGIR, Lars
STEMMANN, Delphine THIBAULT-BOTHA, Céline VIGNOT.
Atelier « Poissons » : Daniela BANARU, Jérôme BAUDRIER, Jean-Louis BIGOT, Philippe
CURY, Romain DAVID, Eric FEUNTEUN, Laurent GUERIN, Laurence LE DIREACH,
François Le LOC’H, Morgane LEJART, Philippe LENFANT, Jérémy PASTOR, Sébastien
PERSONNIC, Gabriel REYGONDEAU, Sandrine VAZ, Héloïse YOU.
Atelier « Prédateurs supérieurs » : Matthieu AUTHIER, Sylvain BONHOMMEAU,
Emmanuel CAILLOT, Léa DAVID, Hélène LABACH, Jean-Paul ROBIN, Jérôme SPITZ,
Aurore STERCKEMAN, Olivier VAN CANNEYT, Cécile VINCENT.
Visioconférence «systèmes haute-résolution de suivi des masses d’eau pélagiques » : Éric
MACE, Isabelle GAILHARD-ROCHER, Alain LEFEBVRE, Felipe ARTIGAS, Pascal
CLAQUIN, Laurent GUERIN, Gérald GREGORI, Melilotus THYSSEN
Autres contributions : Pierrick BOCHER, Tiphaine CHOUVELON, Benoît LEBRETON.
Relecture : Bernard CADIOU, Sylvain LAMARE, Pierre-Guy SAURIAU, Cécile VINCENT.
Liste des abréviations
AAMP : Agence des Aires Marines Protégées
BEE : Bon état écologique
BDC : Comité Biodiversité (OSPAR)
D1 à D11 : Descripteurs, relatifs au bon état écologique de la DCSMM
DCE : Directive Cadre Eau
DCSMM : Directive Cadre « Stratégie pour le Milieu Marin »
EI : Evaluation initiale
GIEC : Groupe Intergouvernemental d’Etude du Climat

OE : Objectifs environnementaux
PAC : Programme d’acquisition de connaissances (DCSMM)
PACOMM : Programme d'Acquisition de Connaissances sur les Oiseaux et les Mammifères Marins en
France métropolitaine
PAMM : Plan d’Action pour le Milieu Marin
PDS : Programme de surveillance (DCSMM)
PVM : Profileur vidéo marin
RNE : Réseau National d’échouages
RNF : Réserves Naturelles de France
T1 à T11 : Thématiques (équivalent des descripteurs), relatives au PDS et PAC de la DCSMM
SRM : Sous-région marine
UVC : Underwater visual census
Pour citer ce document :
DCSMM - Groupe D4, 2013. Programme de surveillance et plan d’acquisition de connaissances pour
la Directive Cadre Stratégie pour le Milieu Marin (DCSMM) : Thématique « Réseaux trophiques ».
(Rapport pour le projet DCSMM).

Sommaire
I. Introduction......................................................................................................................................... 1
I.1. Éléments de contexte généraux sur le thème ................................................................................ 1
I.1.1. Rappels sur les indicateurs du D4.......................................................................................... 2
I.1.2. Groupes biologiques à considérer.......................................................................................... 3
I.2. Structure du document et choix effectués ..................................................................................... 5
I.2.1. Stratégie générale de surveillance pour la thématique 4 ........................................................ 5
II. Objet de la surveillance n°1 : Structure des réseaux trophiques ......................................................... 7
II.1. Paramètres ou groupe de paramètres à suivre ............................................................................. 7
II.2. Objectifs recherchés et enjeux pour la DCSMM ......................................................................... 7
II.2.1. Biomasse, abondance et composition ................................................................................... 7
II.2.2. Répartition des espèces ........................................................................................................ 9
II.2.3. Traits biologiques .............................................................................................................. 10
II.3. Recommandations sur le dispositif de suivi .............................................................................. 11
II.3.1. Méthode recommandée ...................................................................................................... 11
II.3.2. Risques de limites techniques ou opérationnelles à prendre en compte ............................. 30
II.3.3. Recommandations pour la gestion et la bancarisation des données .................................... 31
II.3.4. Recommandations sur les acteurs, les opérateurs potentiels............................................... 31
II.3.5. Informations éventuelles sur les coûts................................................................................ 32
II.3.6. Optimisation intra-thématique ........................................................................................... 33
II.3.7. Optimisation inter-thématique ........................................................................................... 33
II.3.8. Optimisation communautaire ............................................................................................. 34
III. Objet de la surveillance n°2 : Productivité des espèces .................................................................. 35
III.1. Paramètres ou groupes de paramètres à suivre ........................................................................ 36
III.2. Objectifs recherchés et enjeux pour la DCSMM ..................................................................... 36
III.2.1. Production primaire .......................................................................................................... 36
III.2.2. Paramètres liés à la productivité des autres consommateurs ............................................. 37
III.3. Recommandations sur le dispositif de suivi ............................................................................. 38
III.3.1. Méthode recommandée ..................................................................................................... 38
III.3.2. Risques de limites techniques ou opérationnelles à prendre en compte ............................ 42
III.3.3. Recommandations pour la gestion et la bancarisation des données .................................. 43
III.3.4. Recommandations sur les acteurs, les opérateurs potentiels ............................................. 43
III.3.5. Informations éventuelles sur les coûts .............................................................................. 43
III.3.6. Optimisation intra-thématique .......................................................................................... 47
III.3.7. Optimisation inter-thématique .......................................................................................... 47
III.3.8. Optimisation communautaire ............................................................................................ 47
IV. Objet de la surveillance n°3 : Étude des régimes alimentaires et évaluation des niveaux trophiques
............................................................................................................................................................. 49

IV.1. Paramètres ou groupes de paramètres ...................................................................................... 49
IV.2. Objectifs recherchés et enjeux pour la DCSMM ..................................................................... 49
IV.2.1. Zones d’alimentation des prédateurs supérieurs ............................................................... 49
IV.2.2. Valeur énergétique des proies ........................................................................................... 49
IV.2.3. Contenus stomacaux, analyse de fèces et signatures isotopiques (détermination des
niveaux trophiques des espèces et des régimes alimentaires des prédateurs supérieurs) ............... 50
IV.3. Recommandations sur le dispositif de suivi............................................................................. 52
IV.3.1. Méthode recommandée .................................................................................................... 52
IV.3.2. Risques de limites techniques ou opérationnelles à prendre en compte ............................ 64
IV.3.3. Recommandations pour la gestion et la bancarisation des données .................................. 65
IV.3.4. Recommandations sur les acteurs, les opérateurs potentiels ............................................. 65
IV.3.5. Informations éventuelles sur les coûts .............................................................................. 65
IV.3.6. Optimisation intra-thématique .......................................................................................... 82
IV.3.7. Optimisation inter-thématique .......................................................................................... 82
IV.3.8. Optimisation communautaire ........................................................................................... 82
V. Conclusion....................................................................................................................................... 83
V.1. Besoins en termes de paramètres pour la thématique T4 .......................................................... 83
V.2. Besoins en termes de programmes de surveillance pour la thématique T4 ............................... 83
V.3. Besoins en termes d’analyse de données et de bancarisation pour la thématique T4 ................ 86
VI. Références citées ............................................................................................................................ 87
Annexe 1 : Ateliers et réunions ayant contribué aux réflexions en vue de l’élaboration de ce rapport . 93
Annexe 2 : Classification des espèces de poissons en guildes trophiques (CIEM) ............................... 95
Annexe 3 : Fiches techniques des indicateurs communs proposés par le groupe d’experts « Réseaux
trophiques » d’OSPAR/ICG-COBAM (extrait du document BDC 13/4/2 Add.1-E) ............................ 99
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
1
/
140
100%